
ISSN 2472-1972
Effects of Rapid Weight Loss on Systemic and
Adipose Tissue Inflammation and
Metabolism in Obese
Postmenopausal Women
Jos´eO.Alem´an,
1,2,8
Neil M. Iyengar,
2
Jeanne M. Walker,
3
Ginger L. Milne,
4
Joel Correa Da Rosa,
3
Yupu Liang,
3
Dilip D. Giri,
5
Xi Kathy Zhou,
6
Michael N. Pollak,
7
Clifford A. Hudis,
2
Jan L. Breslow,
1
Peter R. Holt,
1
*
and Andrew J. Dannenberg
8
*
1
Laboratory of Biochemical Genetics and Metabolism, Rockefeller University, New York, New York
10065;
2
Department of Medicine, Memorial Sloan-Kettering Cancer Center, New York, New York 10065;
3
Rockefeller Hospital, Rockefeller University, New York, New York 10065;
4
Division of Clinical
Pharmacology, Vanderbilt University Medical Center, Nashville, Tennessee 37232;
5
Department of
Pathology, Memorial Sloan-Kettering Cancer Center, New York, New York 10065;
6
Department of Health
Care Policy and Research, Weill Cornell Medical College, New York, New York 10065;
7
Departments of
Medicine and Oncology, McGill University, Montreal, Quebec H3T 1E2, Canada; and
8
Department of
Medicine, Weill Cornell Medical College, New York, New York 10065
*These authors are co-senior authors.
Context: Obesity is associated with subclinical white adipose tissue inflammation, as defined by the
presence of crown-like structures (CLSs) consisting of dead or dying adipocytes encircled by macro-
phages. In humans, bariatric surgery-induced weight loss leads to a decrease in CLSs, but the effects of
rapid diet-induced weight loss on CLSs and metabolism are unclear.
Objective: To determine the effects of rapid very-low-calorie diet-induced weight loss on CLS density,
systemic biomarkers of inflammation, and metabolism in obese postmenopausal women.
Design: Prospective cohort study.
Setting: Rockefeller University Hospital, New York, NY.
Participants: Ten obese, postmenopausal women with a mean age of 60.6 years (standard deviation, 6
3.6 years).
Main Outcome Measures: Effects on CLS density and gene expression in abdominal subcutaneous
adipose tissue, cardiometabolic risk factors, white blood count, circulating metabolites, and oxidative
stress (urinary isoprostane-M) were measured.
Results: Obese subjects lost approximately 10% body weight over a mean of 46 days. CLS density
increased in subcutaneous adipose tissue without an associated increase in proinflammatory gene
expression. Weight loss was accompanied by decreased fasting blood levels of high-sensitivity C-reactive
protein, glucose, lactate, and kynurenine, and increased circulating levels of free fatty acids, glycerol,
b-hydroxybutyrate, and 25 hydroxyvitamin D. Levels of urinary isoprostane-M declined.
Conclusion: Rapid weight loss stimulated lipolysis and an increase in CLS density in subcutaneous
adipose tissue in association with changes in levels of circulating metabolites, and improved systemic
biomarkers of inflammation and insulin resistance. The observed change in levels of metabolites
Abbreviations: CLS, crown-like structure; CVD, cardiovascular disease; GSEA, Gene Set Enrichment Analysis; GSVA, Gene Set
Variation Analysis; H&E, hematoxylin and eosin; hsCRP, high-sensitivity C-reactive protein; PCR, polymerase chain reaction;
SHBG, steroid hormone-binding globulin; VLCD, very-low-calorie diet.
Received 23 February 2017
Accepted 20 April 2017
First Published Online 25 April 2017
June 2017 | Vol. 1, Iss. 6
doi: 10.1210/js.2017-00020 | Journal of the Endocrine Society | 625–637

(i.e., lactate, b-hydroxybutyrate, 25 hydroxyvitamin D) may contribute to the anti-inflammatory effect
of rapid weight loss.
Copyright © 2017 Endocrine Society
This article has been published under the terms of the Creative Commons Attribution Non-
Commercial, No-Derivatives License (CC BY-NC-ND; https://creativecommons.org/licenses/by-nc-
nd/4.0/).
Freeform/Key Words: crown-like structure, weight loss, metabolism, inflammation
Obesity has reached epidemic proportions, with about two-thirds of US adults classified as
overweight or obese [1]. Complications of obesity include type 2 diabetes, cardiovascular
disease, and increased incidence of and worsened prognosis for several cancers, including
postmenopausal breast cancer [2, 3]. Inflamed adipose tissue is believed to contribute to
obesity-associated complications. This is characterized by immune-cell infiltration, increased
levels of proinflammatory cytokines, as well as adipocyte insulin resistance, mitochondrial
dysfunction, and endoplasmic reticulum stress [4]. Crown-like structures (CLSs) are found in
the white adipose tissue of both obese mice and humans. In cross-sectional studies, the
presence of CLSs is associated with ins ulin resistance, cardiovascular disease (CVD), and
worse prognosis for patients with cancer [ 5–7]. These inflammatory foci represent dead or
dying adipocytes enveloped by macrophages. The macrophages rely on lysosomal exophagy to
phagocytose the dead adipocytes and become foam cells [8, 9]. When white adipose tissue fails
to expand appropriately to store excess energy, ectopic fat deposition occurs in other organs,
leading to insulin resistance—a process known as lipoto xicity. The ability of the CLS mac-
rophages to store free fatty acids may help protect against both local adipocyte dysfunction
and lipotoxicity [10].
In obese rodents, prolonged caloric restriction is associated with a reduction in CLS and
reduced expression of proinflammatory genes [11]. By contrast, rapid weight-loss results in
increased macrophage infiltration in visceral and subcutaneous adipose tissue [12]. In
humans, bariatric surgery-induc ed weight loss leads to a reduction in CLSs, improved
systemic inflammation, and decreased insulin resistance and, possibly, cancer risk [ 13]. The
effects of very-low-calorie diet (VLCD)-mediated rapid weight loss have also been in-
vestigated. Cl´ement and colleagues [14] reported that VLCD-induced rapid weig ht loss leads
to a substantial reductio n in fat mass in association with improved insulin sensitivity, re-
duced circulating triglyceride and cholesterol levels, as well as changes in subcutaneous fat
gene expression. There is considerable evidence that cross-talk between adipose tissue and
distal tissues, including the liver, pancreas, and skeletal muscle, can affect health [15].
Although the effects of rapid weight loss on circulating levels of adipokines and lipids have
been reported [16, 17], much less is known about the effects on systemic levels of metabolites.
In this study, our primary objective was to investigate the effects of VLCD-induced weight
loss on CLS den sity, systemic biomarkers of inflammation, and metabolism in obese post-
menopausal women.
1. Materials and Methods
A. Subjects
Eligible subjects were obese (body mass index, $35 kg/m
2
) postmenopausal women (defined
by .2 years without menstrual periods). Participants were recruited from the community
through advertis ing or participation in a Rockefeller Hospital registry of subjects previously
screened for research studies. We excluded subjects with clinical CVD; type 2 diabetes and
receiving oral hypoglycemic agents; smokers; regular users of aspirin, fish oils, or vitamin D
supplements; a history of gastrointestinal surgery other than appendectomy; or breast and
other cancers. Fourteen subjects were screened, and 12 of these were evaluated and enrolled.
During the study, two subjects dropped out: one because of acute conjunctivitis and another
626 | Journal of the Endocrine Society | doi: 10.1210/js.2017-00020
for inability to adhere to the diet. Of those who completed the study, three were white, four
were bl ack, and three were Hispanic. This was a single-center study performed at The
Rockefeller University Hospital between September 2012 and August 2013. Subjects
underwent a complete medical examination, standard blood and urine tests, and an elec-
trocardiogram, and all were found to be healthy. A Transparent Reporting of Evaluation with
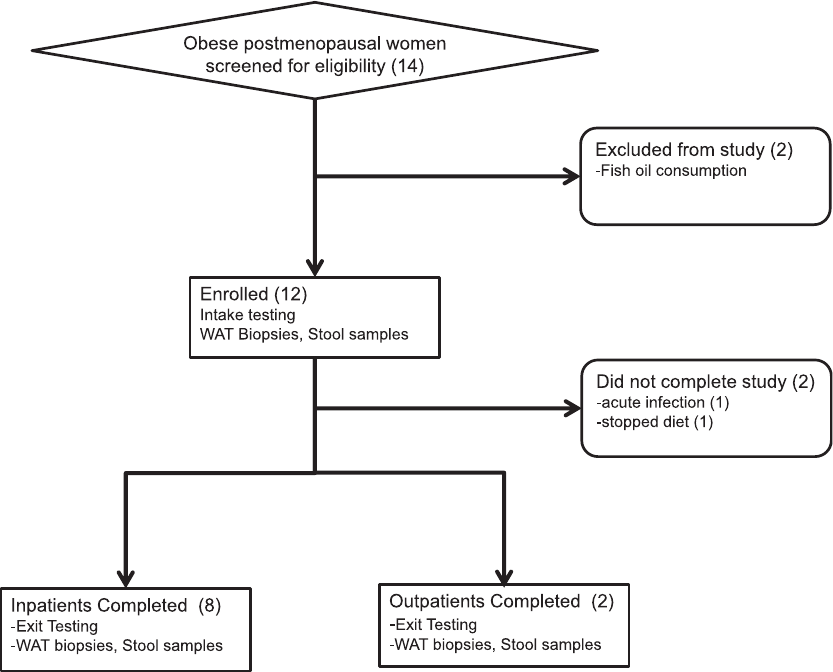
Nonrandomized Designs flowchart summ arizing subject selection is presented in Figure 1.
B. Sample Size
The primary end point of this study was to determine whether the intervention would sig-
nificantly alter CLS density in subcutan eous adipose tissue. Sample size was primarily
determined by logistics and feasibility considerations. However, with 10 subjects, assuming a
coefficient of variation of 30%, a paired t test applied to this sample provided 80% power to
detect an effect size as small as a 1-unit change in CLS/cm
2
from before to after weight loss.
This effect size is equivalent to 30% increase from baseline CLS density.
C. Trial Approval and Registration
The study was approved by the institutional review boards at Rockefeller University, Weill
Cornell Medical College, and Memorial Sloan-Kettering Cancer Center, and registered under
ClinicalTrials.gov identifier NCT01699906.
Figure 1. Transparent Reporting of Evaluation with Nonrandomized Designs flowchart of
the study. Fourteen subjects were screened, of whom 12 were enrolled, and 10 completed the
study. Two subjects completed the weight loss intervention as outpatients, and the remaining
subjects completed as inpatients. WAT, white adipose tissue.
doi: 10.1210/js.2017-00020 | Journal of the Endocrine Society | 627
D. Interventions, Specimen Collect ion, and Analysis
The effect of the VLCD was assessed in subjects after they had lost approximately 10% of body
weight. The diet of each subject before weight loss was evaluated by the Harvard Food
Frequency Questionnaire, which quantifies food intake over a 3-day period [18]. On admis-
sion to the Rockefeller University Hospital, baseline study specimens were collected. Sub-
sequently, participants underwent a 3-day dietary adjustment period, consuming 50% of their
prestudy caloric intake. Of the 10 subjects who completed the VLCD-induced weight loss, two
did so as outpatients with intensive monitoring and eight as inpatients. Blood samples were
drawn biweekly for safety monitoring, which included measurements of electrolytes and liver
function tests. The rate of weight loss did not differ significantly between those studied as
outpatients or inpatients. After losing approximately 10% of their baseline body weight, all
subjects consumed the VLCD for three additional days as inpatients, during which all of the
baseline measurements were repeated. Bloods for end-point analyses were collected under
fasting conditions. Therefore, each study subject acted as her own control.
The VLCD consisted of a commercially available diet (New Direction Program, Robard
Corp., Mount Laurel, NJ) that provided approximately 800 Kcal/d with an estimated mac-
ronutrient energy distribution of 54% protein, 26% carbohydrate, 20% fat (including 4%
saturated fat and 200 mg of cholesterol) and 10 g/d fiber, as previou sly described [19]. This
commercial preparation provided a choice of shakes, soups, bars, and puddings. Subjects had
four choices per day and consumed one item every four waking hours.
Fasting blood samples were analyzed in the Clinical Pathology Laboratory of Memorial
Sloan-Kettering Cancer Center for electrolyte levels, liver function, renal function, lipid
profile, and high-sensitivity C-reactive protein (hsCRP) level. Aliquots of serum for cytokine
measurements, including interleukin (IL)-6, IL-10, and IL-17, were stored at 280°C for
subsequent analysis. Enzyme-linked immunosorbent analyses of leptin, adiponectin, insulin,
steroid hormone-binding globulin (SHBG), estradiol, and IL-6 were performed at the Pollak
Assay Laboratory (McGill University, Quebec , Canada) [6]. Erenna immunoassay analyses
(EMD Millipore, Billerica, MA) of IL-10 and IL-17 were performed by the Rockefeller
Translational Technology Core Laboratory. Body composition was measured by air-
displacement plethysm ography using the BodPod system (COSMED, Italy). Subcutaneous
adipose tissue was obtained by aspiration biopsy using a large-bore, modified liposuction
needle (Anthony Products, Indianapolis, IN) under local anesthesia with lidocaine between
08:00 and 11:00. The initial adipose tissue biopsy specimen was taken in the left lower
quadrant of the abdomen of each subject, whereas the final biopsy specimen was taken in the
right lower quadrant abdomen. The quadrants for the two biopsies were alternated to avoid
the potential of results being confounded by trauma-related to the initial biopsy. Adipose
tissue was formalin fixed and paraffin embedded for immunohistochemistry or frozen in
RNAlater (Ambion Inc, Austin TX) for subsequent RNA extraction and analysis.
E. Urinary F2-isoprostane-M Measurement
Analysis of urinary F2-isoprostane-M was performed on an aliquot of a 24-hour urine col-
lection by liquid chromatography–mass spectrometry in the Vanderbilt University Eicosa-
noid Core Laboratory as described previously [20]. F2-isoprostane-M levels were normalized
to creatinine.
F. CLS Quantification in Adipose Tissue Biopsy Specimens
CLS in abdominal subcutaneous adipose tissue bi opsy specimens were quantified as de-
scribed previously [6]. Biopsy samples were fixed in formalin for 24 hours and then embedded
in a paraffin block. Six sections were cut with a spacing of 50 mm; the first section was stained
with hematoxylin and eosin (H&E) to ensure the sample was representative of adipose tissue.
The remaining five sections were stained for the macrophage marker CD68 (mouse mono-
clonal KP1 antibody; dilution 1:4000; Dako, Glostrup, Denmark) to identify CLS by light
628 | Journal of the Endocrine Society | doi: 10.1210/js.2017-00020
microscopy. The numbers of CLSs per section were recorded by the study pathologist (D.D.G.).
The area of adipose tissue examined in each of the five sections was measured using Image J
software (National Institutes of Health, Bethesda, MD) to calculate CLS density as CLS/cm
2
.
G. Adipocyte Diameter
H&E-stained sections were generated from subcutaneous adipose tissue biopsy specimens to
measure adipocyte diameters, as previously described [21, 22]. The H&E-stained sections
were photographed at 320 magnification using an Olympus BX50 microscope (Orangeburg,
NY) and MicroFire digital camera (Optronics Goleta, CA). Mean diamet ers were calcu-
lated using measurements from $30 individual adipocytes for each patient, using the linear
dimensional tool in the Canvas 11 Software (ACD Systems International Victoria, Canada).
H. Gene Expression
Total RNA was extracted from approximately 0.5 g of frozen adipose tissue using a Qiagen
RNeasy Lipid Tissue Mini Kit (Germantow n, MD). RNA quality was assessed using an
Agilent Bioanalyzer. Approximately, 2 to 3 mg of RNA with RNA integrity number .7 were
submitted for 50-bp paired-end read RNA sequencing of polyA-enriched RNA at the New York
Genome Center. Gene expression was analyzed for pathway enrichment using Gene Set
Enrichment Analysis (GSEA; Broad Institute, Cambridge MA) and Ingenuity Pathway
Analysis (Qiagen).
For heat map generation, genes were annotated using the biomaRt library in R package
Bioconductor (https://www.bioconductor.org/). RNA-seq data were normalized by a regu-
larized logarithmic transformation into a ma trix through DESeq2 [23]. The Gene Set Var-
iation Analysis (GSVA) algorithm [24] was applied to the matrix to quantify pathway activity
for a set of 1454 curated gene sets available in the Molecular Signatures Database. Supervised
heat maps were generated to visually compare pathway activities in before and after weig ht-
loss samples, and to assess subject and pathway similarities. A linear mixed-effects model
was applied to estimate averages for the pathway activity before and after weight loss and to
determine the pathways that differed significantly between before and after VLCD weight
loss. To achieve a more concise visual representation, these averages were displayed in a heat
map (scaled column-wise) based on Euclidean distance and a complete linkage algorithm for
pathway hierarchical cluster ing.
I. Quantitative Real-Time Polymerase Chain Reaction
Transcript expression was assessed after total RNA was isolated from adipose tissue, using
the RNeasy Mini Kit (Qiagen). RNA (100 ng) was reverse transcribed using the qScript cDNA
Synthesis Kit (QuantaBio Beverly, MA) and the resulting cDNA used for real-time poly-
merase chain reaction (PCR) amplif ication using Fast SYBR Green PCR master mix on a 7500
HT real-time PCR system (Applied Biosystems, Foster City, CA). IL-6 was amplified using a
Qiagen QuantiTect primer assay (catalog no. QT00083720). Primers used to amplify tumor
necrosis factor-a were: forward: 5
0
-CTG CAC TTT GGA GTG AT-3
0
; reverse: 5
0
-AGA TGA TCT
GAC TGC CTG GG-3
0
. Primers used to amplify IL-1b were: forward: 5
0
-GGA CAA GCT GAG
GAA GAT GC-3
0
; reverse: 5
0
-TCG TTA TCC CAT GTG TCG AA-3
0
. Transcript expression was
normalized to glycerald ehyde 3-phosphate dehydrogenase (GAPDH), which was amplified
using a Qiagen QuantiTect primer assay (catalog no. QT00079247). Expression was de-
termined using the DDCT analysis protocol.
J. Statistical Analyses
Two-tailed paired t tests and Wilcoxon tests were used to compare anthropometric mea-
surements, biochemical variables, serum cytokines, and immunohistochemical results before
and after VLCD-induced weight loss. P , 0.05 was considered significant. For metabolomic
doi: 10.1210/js.2017-00020 | Journal of the Endocrine Society | 629

analyses, matched paired t tests were used to identify compounds that differed sign ificantly
between the before and after weight-loss samples, correcting for multiple measurements.
GraphPad Prism and Excel were used for data visualization.
2. Results
The mean age (6 standard dev iation) of the 10 subjects completing the study was 60.6 6
3.6 years. The mean time to induce ~10% weight loss was 46.2 6 15.3 days (range, 30 to 74 days).
Table 1 shows the mean macronutrient composition of the subjects’ self-selected prestudy
diets and that of the VLCD consumed during weight loss. The daily protein intake of the
VLCD (388 6 45 Kcal protein per day) was similar to the subject’sprestudydiet(3826 28 Kcal
protein per day; P = not significant), whereas fat, carbohydrate, and fiber contents were all
markedly reduced.
A. Effects of the VLCD-Induced Weight Loss on Anthropometric and Clinical Parameters
The before and after weight loss anthropometric and clinical data are shown in Table 2. The
VLCD decreased BMI by 9.6%, including a 3.8% reduction in body fat. Decreases were ob-
served in fasting glucose (13.3%) and insulin (20.3%) levels, and the Homeostatic Model for
Assessment of Insulin Resistance (30.9%), reflecting improved insulin sensitivity. High-
density lipoprotein cholesterol levels decreased (16.0%), with trends towar d decreased
low-density lipoprotein cholesterol (8.2%) and triglyceride (19.3%) levels. Fasting leptin levels
decreased (57.2%), whereas adiponectin was unaffected. Estradiol levels were unaffected by
weight loss, but levels of SHBG increased (56.7%). Weight loss led to a sign ificant increase in
25-hydroxyvitamin D (19.6%) and calcium (2.5%) levels.
B. Effects of the VLCD-Induced Weight Loss on Systemic Markers of Inflammation and
Oxidant Stress
VLCD-induced weight loss was associated with decreased levels of hsCRP (35.3%) and total
white blood cell count (17.4%), largely due to fewer circulating neutrophils (28.3%) (Table 2).
Levels of urinary F2 isoprostane-M, a biomarker of oxidant stress, decreased by 41.1% fol-
lowing VLCD-induced weight loss.
C. Effects of the VLCD-Induced Weight Loss on Subcutaneous Adipose Tissue
Mean adipocyte diameter was 111.1 6 13.0 mm at baseline and 111.7 6 11.3 mm after weight
loss. We also assessed the density of CLSs in abdominal subcu taneous adipose tissue at
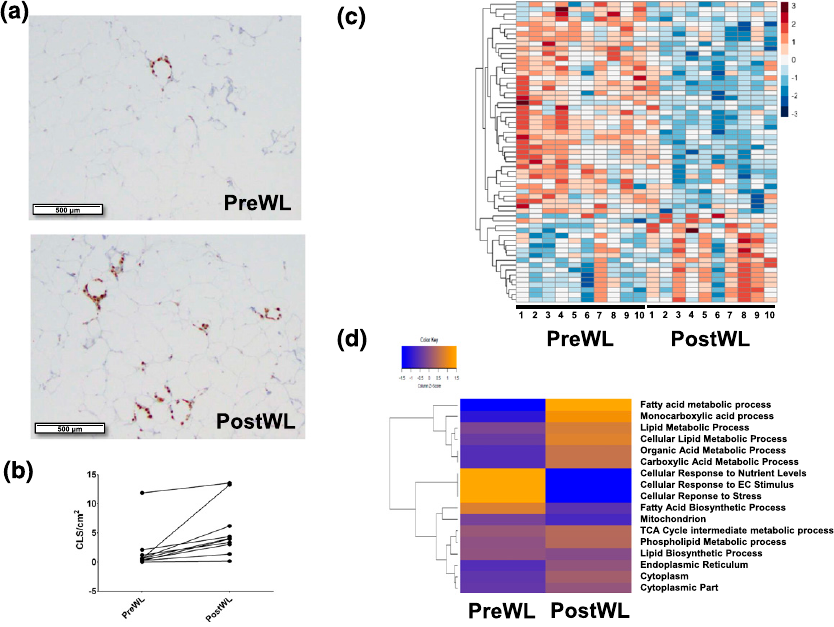
baseline and after weight loss [ Fig. 2(a)]. CLSs were detected in all 10 subjects at bas eline.
Interestingly, the density of CLSs increa sed from a mean of 1.78 6 1.04 to 5.33 61.32 CLS/cm
2
after weight loss [P = 0.01; Fig. 2(b)].
Table 1. Composition of the Diet Before Weight Loss and the VLCD
preWL Diet VLCD
Calories, Kcal/d 2324 6 1041 721 6 83
% Carbohydrate 49 6 8266 1
% Protein 16 6 3546 1
% Fat 36 6 8206 2
% Saturated fat 11 6 446 1
Fiber, g 33 6 18 10 6 4
Values are presented as mean 6 standard deviation.
Abbreviation: preWL, before weight loss.
630 | Journal of the Endocrine Society | doi: 10.1210/js.2017-00020

To further explore the effects of VLCD-induced weight loss on the biology of subcutaneous
adipose tissue, RNA sequencing was carried out on adipose tissue obtained before and after
weight loss. A total of 61 genes were differentially expressed at absolute log fold-change .2
and P , 0.001 (Supplemental Table 1). Of the differentially express ed genes, about two-thirds
decreased and one-third increased after weight loss [Fig. 2(c)]. GSEA identified over-
represented metabolic pathways relating to triacylglycerol and ketone metabolism, and
cellular respiration. As shown in Figure 2(d), analysis by GSVA showe d that VLCD-induced
weight loss was associated with increased expression of genes involved in metabolism of
monocarboxylic acids, fatty acids, the tricarboxylic acid cycle, phospholipids, lipids, organic
acid, and carboxylic acids; and decreas ed expression of genes involved in fatty acid bio-
synthesis, cellular responses to nutrient levels, extracellular signaling, and to stress. Ad-
ditionally, pathway analyses, including GSEA, Ingenuity Pathway Analysis, and GSVA
showed that VLCD-induced weight loss did not affect the expression of genes related to
inflammation. To confirm this finding, quantitative real-time PCR was carried out. Levels of
tumor necrosis factor-a, IL-1b, and IL-6 mRNAs were unaffected by VLCD-induced weight
loss (data not shown).
D. Effects of VLCD-Induced Weight Loss on the Plasma Metabolome
To determine the effects of VLCD-induced weight loss on circulating metabolites, plasma
collected under fasting conditions was subjected to targeted metabolomic analysis. A total of
336 metabolites were detected. Of these, 131 metabolites changed significantly, with 67
Table 2. Effects of Rapid VLCD-Induced Weight Loss
preWL postWL Difference, % P Value
a
Weight, kg 101.3 6 9.7 90.9 6 8.2 210.3 ,0.01
BMI, kg/m
2
38.8 6 3.4 35.1 6 3.0 29.6 0.01
Waist, cm 115.8 6 6.3 107.5 6 7.9 27.2 0.01
% Body fat composition 51.4 6 6.1 47.6 6 5.5 27.4 ,0.01
Metabolic parameters
Glucose, mg/dL 103.4 6 17.6 89.6 6 14.9 213.3 0.01
Insulin, mU/mL 12.3 6 1.8 9.8 6 2.1 220.3 0.07
HOMA-IR 3.30 6 0.59 2.28 6 0.60 230.9 0.04
HDL, mg/dL 54.3 6 10.3 45.6 6 6.2 216.0 0.02
LDL, mg/dL 131.2 6 24.8 120.5 6 27.0 28.2 0.08
TG, mg/dL 96.9 6 28.8 78.2 6 25.0 219.3 0.08
Leptin, ng/mL 61.0 6 21.0 26.1 6 12.2 257.2 ,0.001
Adiponectin, mg/mL 9.8 6 5.8 10.2 6 5.5 4.1 0.20
Estradiol, pg/mL 36.2 6 14.3 33.4 6 11.6 27.7 0.38
SHBG, nmol/L 60.3 6 26.3 94.5 6 33.7 56.7 ,0.001
Calcium, mg/dL 9.06 6 0.27 9.29 6 0.31 2.5 0.01
25-OH vitamin D, ng/mL 22.5 6 10.3 26.9 6 10.3 19.6 0.04
Inflammatory parameters
WBC, 10
3
/uL 5.96 6 2.00 4.92 6 1.56 217.4 0.01
Neutrophils, 10
3
/mL 3.22 6 0.60 2.31 6 0.54 228.3 0.01
hsCRP, mg/dL 0.68 6 0.42 0.44 6 0.24 235.3 0.01
IL-6, pg/mL 3.1 6 1.7 2.6 6 1.1 216.1 0.07
IL-10, pg/mL 1.49 6 0.99 1.31 6 0.76 212.1 0.50
IL-17, pg/mL 0.22 6 0.10 0.24 6 0.16 9.1 0.38
MCP-1, pg/mL 312 6 130 292 6 115 26.4 0.32
F2-IsoP-M, ng/mg Cr 1.51 6 0.57 0.89 6 0.29 241.1 0.01
Values are presented as mean 6 standard deviation.
Abbreviations: 25-OH vitamin D, 25 hydroxyvitamin D; BMI, body mass index; F2-IsoP-M, F2-isoprostane-M; HDL,
high-density lipoprotein; HOMA-IR, Homeostatic Model for Assessment of Insulin Resistance; LDL, low-density
lipoprotein; MCP-1, monocyte chemoattractant protein 1; TG, triglyceride; WBC, white blood cell count.
a
Significance was determined by Wilcoxon signed-rank test.
doi: 10.1210/js.2017-00020 | Journal of the Endocrine Society | 631
increasing (Supplemental Table 2) and 64 decreasing (Supplemental Table 3). The decrease in
plasma levels of creatine and 3-methylhistidine after VLCD-induced weight loss is consistent
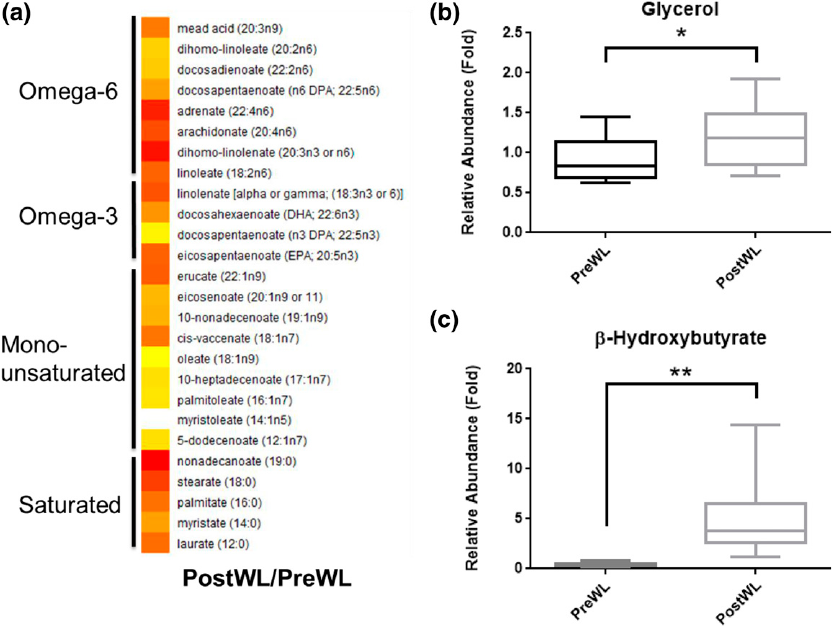
with this diet being protein sparing (Table 3). The observed increase in both circulating free
fatty acid and glycerol levels after treatment with the VLCD suggests enhanced lipolysis [Fig.
3(a) and 3(b)]. Treatment with the VLCD also led to elevated ketone levels, including a 16-fold
increase in b-hydroxybutyrate and a 7.6-fold increase in acetoacetate (Table 3). Levels of
lactate, tryptophan, and the tryptophan metabolite kynurenine all decreased after VLCD-
induced weight loss. By contrast, the metabolites p -cresol sulfate and glycolithocholate
sulfate increased after VLCD-induced weight loss, consistent with altered intestinal bac terial
metabolism (Table 3).
3. Discussion
In this study, we determined the effects of VLCD-induced rapid weight loss on both in-
flammation and metabolism in obese postmenopausal women. Consistent with the known
anti-inflammatory effects of weight loss [25], VLCD-induced rapid weight loss resulted in
decreased levels of hsCRP, reduced white blood cell count, and a near-significant reduction in
Figure 2. Effects of VLCD-induced weight loss on subcutaneous adipose tissue. Biopsy
specimens were obtained before and after weight loss and subjected to CD68 immunohistochemistry
and RNA sequencing. (a) Subcutaneous adipose tissue images from the subject with the
largest increase in CLSs after weight loss are shown. (Top panel) preWL. (Bottom panel)
postWL. (b) Line plot of CLS density (CLS/cm
2
) shown from preWL to postWL in each
individual subject (P = 0.01). (c) Heat map for 61 genes whose expression changed
significantly from preWL to postWL. Subjects (columns) are arranged from preWL to postWL.
Genes (rows) are clustered hierarchically in a supervised manner. (d) Heat map of expression
of 17 lipid metabolism gene pathways altered significantly between preWL and postWL.
postWL, after weight loss; preWL, before weight loss.
632 | Journal of the Endocrine Society | doi: 10.1210/js.2017-00020

circulating IL-6 levels [26]. Obesity has been associated with oxidant stress, which can also
contribute to inflammat ion [27]. We observed decreased levels of urinary F2-isoprostane-M
after weight loss, suggesting decreased oxidative stress [28].
The density of CLS, a histological marker of adipose inflammation, is increased in both
visceral and subcutaneous adipose tissue from obese vs lean subjects and correlates with
elevated levels of circulating proinflammatory mediators [29]. To complement our analysis of
systemic markers of inflammation, we explored the effects of rapid VLCD-induced weight loss
on CLS density in subcutaneous adipose tissue. Interestingly, VLCD-induced weight loss was
associated with a significant increase in CLS density in the absence of a change in adipocyte
size or expression of proinflammatory genes. Weight loss after bariatric surgery has been
associated with a reduction in CLS [13]. By contrast, in obese mice, Kosteli et al. [12] showed
that rapid weight loss stimulated lipolysis, resulting in the accumulation of macrophages in
adipose tissue [12]. Elevated level s of free fatty acids, a consequence of increased lipolysis,
were found to function as chemoattractants, leading to increased macrophage content. We
also observed a significant increase in lipolysis after VLCD-induced weight loss. Taken to-
gether, we speculate that VLCD-mediated rapid weight loss stimulated lipolysis, contributing
to the observed increase in CLS density.
The increase in CLS density in response to VLCD-induced weight loss has potentially
significant implications. These CLS macrophages can store excess free fatty acids [29, 30],
thereby protecting against lipotoxicity and contributing to the observed improvement in
insulin sensitivity and decreased systemic inflammation following rapid weight loss. There
are other possible explanations for how rapid VLCD-induced weight loss could lead to both an
increase in CLS density in abdominal subcutaneous fat and a decrease in systemic in-
flammatory biomarkers. One possibility is that rapid weight loss has different effects on
visceral vs subcutaneous fat. Perhaps a decrease in CLS density occurred in visceral fat,
contributing to reduced systemic inflammation. Another possibility is that the overall de-
crease in fat mass mediated by the VLCD led to a decrease in systemic inflammation. A
limitation of our study is that the effects of rapid weight loss were only measured at a sing le
time point when fat remodeling was occurring. It is possible that a decrease in CLS density
would be observed later, once a new steady state is achieved.
Although weight loss was associated with a small increase in CLS density, we did not
detect a comparable increase in the express ion of proinflammatory genes. There are multipl e
potential explanations for this findin g. Lactate has been observed to stimulate M1 polari-
zation of macrophages [31]. Plasma lactate levels were reduced after rapid weight loss, pre-
sumably reflecting improved adipose tissue oxygenation [32]. Because lactate has been reported
to stimulate M1 polarization of macrophages, a fall in adipose tissue lactate levels in concert with
reduced circulating levels could prevent an increase in proinflammatory gene expression. We
also observed a 16-fold increase in b-hydroxybutyrate after weight loss. Youm et al. [33] found
that b-hydroxybutyrate blocked inflammation mediated by the NLRP3 inflammasome. It is
possible, therefore, that increased b-hydroxybutyrate levels contributed to the lack of a local
inflammatory phenotype in subcutaneous adipose tissue. Consistent with findings in other
weight-loss studies [34], we observed an increase in circulating 25 hydroxyvitamin D levels after
Table 3. VLCD-Induced Weight Loss Was Associated With Changes in Levels of Plasma Metabolites
Increased Plasma
Metabolite
Mean Pairs Ratio
PostWL/PreWL P Value
Decreased Plasma
Metabolite
Mean Pairs Ratio
PostWL/PreWL I Value
b-Hydroxybutyrate 15.98 ,0.001 3-Methylhistidine 0.63 0.01
p-Cresol sulfate 10.05 0.03 Creatine 0.69 ,0.01
Glycolithocholate sulfate 8.76 0.03 Kynurenine 0.74 ,0.001
Acetoacetate 7.64 ,0.001 Lactate 0.74 ,0.01
Tryptophan 0.78 ,0.001
Values are presented as mean pairs ratio of postWL vs preWL levels.
Abbreviations: PostWL, after weight loss; PreWL, before weight loss.
doi: 10.1210/js.2017-00020 | Journal of the Endocrine Society | 633
VLCD-induced weight loss. Because vitamin D possesses anti-inflammatory properties [35], the
increase in 25 hydroxyvitamin D levels might also help to explain the lack of increased
proinflammatory gene expression after VLCD-induced weight loss.
We also determined the effects of VLCD-induced weight loss on a variety of metabolic end
points. Insulin sensitivity improved, as shown by a reduced Homeostatic Model for As-
sessment of Insulin Resistance index. The VLCD diet was associated with reduced levels of
both high-density lipoprotein cholesterol and triglycerides, which, together with a trend
toward decreased low-density lipoprotein cholesterol levels, probably makes the changes
neutral with respect to risk of CVD [36]. Estradiol levels were low, reflecting the post-
menopausal state, and did not change after VLCD-induced weight loss. However, SHBG
levels increased after treatment with the VLCD, which would be predicted to decrease free
estradiol. In all likelihood, the increase in SHBG levels after VLCD-induced weight loss
reflects improved insulin sensitivity [37]. Consistent with the observed decrease in fat mass,
levels of leptin, a proinflammatory adipok ine, declin ed by .50% after VLCD-induced weight
loss. This finding is consistent with the recent report of Magkos et al. [38] who showed that
5.1% to 16.4% weight loss led to a progressive decline in circula ting leptin levels. Increased
circulating levels of estradiol, insulin resistance, and elevated levels of leptin have been
suggested to contribute to the increased risk of breast cancer in obese postmenopausal women
[39]. The changes in free estrogens, insulin resistance, and leptin levels after rapid VLCD-
induced weight loss collectively may lower the risk of breast cancer in this population [40].
Figure 3. Effects of VLCD-induced weight loss on levels of free fatty acids and glycerol in
plasma. (a) Heat map comparing the ratios of plasma free fatty acid levels between postWL
and preWL samples. White indicates unity, with yellow to red indicating increased levels. All
free fatty acid species shown were significantly increased (P , 0.05). (b) Box plot of preWL
and postWL abundance of plasma glycerol relative to total ion counts, and scaled by setting
the preWL loss median to one (*P = 0.01). postWL, after weight loss; preWL, before weight
loss.
634 | Journal of the Endocrine Society | doi: 10.1210/js.2017-00020

Obesity is also associated with numerous immunosuppressive effects [41]. Kynurenine, an
endogenous ligand of the aryl hydrocarbon receptor, has well-documented immunomodu-
latory effects, including stim ulating na
¨
ıve T-cell polarization to the regulatory T-cell phe-
notype [42]. VLCD-induced weight loss was accompanied by reductions in circulating level s of
tryptophan and kynurenine, a tryptophan metabolite. Based on these metabolic changes,
future studies are warranted to evaluate the effects of rapid weight loss on immune function.
In summary, we showed in obese postmenopausal women that rapid diet-induced weight
loss was associated with inc reased CLS density with concomitant improved insulin sensitivity
and decreased circulating biomarkers of inflammation. The increase in CLS density is likely
to reflect adipose remod eling. In cross-sectional studies, we and others have shown that
obesity-associated CLSs are associated with diabetes, CVD, and worse cancer prognosis. The
current study illustrates the dynamic nat ure of CLS and highlights the potential importance
of the rate of weight loss on adipose tissue biology.
Acknowledgments
We thank the clinical research staff and research volunteers who made this work possible. We ac-
knowledge Juana Gonzalez and the Translational Technology Core Laboratory at Rockefeller Univer-
sity Hospital for IL-10 and IL-17 measurements.
Current affiliation: J.O. Alem ´an’s current affiliation is Division of Endocrinology, Diabetes and
Metabolism, Department of Medicine, NYU Langone Medical Center, 522 First Ave, New York, NY
10016. C.A. Huddis’ current affiliation is American Society of Clinical Oncology, 2318 Mill Rd, Suite 800,
Alexandria, VA 22314.
Address all correspondence to: Jos´e Orlando Alem ´an, Rockefeller University, 1230 York Ave, Box 179,
This work was supported by the National Center for Advancing Translational Sciences Grant UL1
TR000043, National Institutes of Health Clinical and Translational Science Award program to
Rockefeller University, the Rockefeller University Center for Basic and Translational Research on
Disorders of the Digestive System, from the Leona M. and Harry B. Helmsley Charitable Trust, The
Rockefeller University Sackler Center for Biomedicine and Nutrition from La Fondation Sackler, the
Breast Cancer Research Foundation, the Botwinick-Wolfensohn Foundation (in memory of Mr. and Mrs.
Benjamin Botwinick), and the Memorial Sloan-Kettering Cancer Center Core Grant (P30 CA008748).
Author contributions: J.O. A., N.M.I., C.A.H., J.L.B., P.R.H., and A.J.D. designed the clinical study.
J.O.A. and J.M.W. collected clinical study end points. J.O.A., N.M.I., J.M.W., G.L.M., J.C.R., Y.L., D.D.G.,
X.K.Z., and M.N.P. analyzed the data. J.O.A., J.L.B., P.R.H., and A.J.D. wrote the manuscript. J.L.B. and
A.J.D. edited the manuscript.
Clinical trial registry: ClinicalTrials.gov no. NCT01699906 (registered 12 September 2012).
Disclosure summary: The authors have nothing to disclose.
References and Notes
1. Institute of Medicine. Accelerating Progress in Obesity Prevention: Solving the Weight of the Nation.
Washington, DC: National Academies Press; 2012.
2. van Kruijsdijk RC, van der Wall E, Visseren FL. Obesity and cancer: the role of dysfunctional adipose
tissue. Cancer Epidemiol Biomarkers Prev. 2009;18(10):2569–2578.
3. Alem ´an JO, Eusebi LH, Ricciardiello L, Patidar K, Sanyal AJ, Holt PR. Mechanisms of obesity-induced
gastrointestinal neoplasia. Gastroenterology. 2014;146(2):357–373.
4. Qatanani M, Lazar MA. Mechanisms of obesity-associated insulin resistance: many choices on the
menu. Genes Dev. 2007;21(12):1443–1455.
5. Apovian CM, Bigornia S, Mott M, Meyers MR, Ulloor J, Gagua M, McDonnell M, Hess D, Joseph L,
Gokce N. Adipose macrophage infiltration is associated with insulin resistance and vascular endothelial
dysfunction in obese subjects. Arterioscler Thromb Vasc Biol. 2008;28(9):1654–1659.
6. Iyengar NM, Zhou XK, Gucalp A, Morris PG, Howe LR, Giri DD, Morrow M, Wang H, Pollak M, Jones
LW, Hudis CA, Dannenberg AJ. Systemic correlates of white adipose tissue inflammation in early-stage
breast cancer. Clin Cancer Res. 2016;22(9):2283–2289.
doi: 10.1210/js.2017-00020 | Journal of the Endocrine Society | 635
7. Iyengar NM, Ghossein RA, Morris LG, Zhou XK, Kochhar A, Morris PG, Pfister DG, Patel SG, Boyle JO,
Hudis CA, Dannenberg AJ. White adipose tissue inflammation and cancer-specific survival in patients
with squamous cell carcinoma of the oral tongue. Cancer. 2016;122(24):3794–3802.
8. Xu X, Grijalva A, Skowronski A, van Eijk M, Serlie MJ, Ferrante AW, Jr. Obesity activates a program of
lysosomal-dependent lipid metabolism in adipose tissue macrophages independently of classic acti-
vation. Cell Metab. 2013;18(6):816–830.
9. Haka AS, Barbosa-Lorenzi VC, Lee HJ, Falcone DJ, Hudis CA, Dannenberg AJ, Maxfield FR. Exo-
cytosis of macrophage lysosomes leads to digestion of apoptotic adipocytes and foam cell formation.
J Lipid Res. 2016;57(6):980–992.
10. Cinti S, Mitchell G, Barbatelli G, Murano I, Ceresi E, Faloia E, Wang S, Fortier M, Greenberg AS, Obin
MS. Adipocyte death defines macrophage localization and function in adipose tissue of obese mice and
humans. J Lipid Res. 2005;46(11):2347–2355.
11. Bhardwaj P, Du B, Zhou XK, Sue E, Harbus MD, Falcone DJ, Giri D, Hudis CA, Kopelovich L,
Subbaramaiah K, Dannenberg AJ. Caloric restriction reverses obesity-induced mammary gland in-
flammation in mice. Cancer Prev Res (Phila). 2013;6(4):282–289.
12. Kosteli A, Sugaru E, Haemmerle G, Martin JF, Lei J, Zechner R, Ferrante AW, Jr. Weight loss and
lipolysis promote a dynamic immune response in murine adipose tissue. J Clin Invest. 2010;120(10):
3466–3479.
13. Cancello R, Henegar C, Viguerie N, Taleb S, Poitou C, Rouault C, Coupaye M, Pelloux V, Hugol D,
Bouillot JL, Bouloumi´e A, Barbatelli G, Cinti S, Svensson PA, Barsh GS, Zucker JD, Basdevant A,
Langin D, Cl´ement K. Reduction of macrophage infiltration and chemoattractant gene expression
changes in white adipose tissue of morbidly obese subjects after surgery-induced weight loss. Diabetes.
2005;54(8):2277–2286.
14. Cl´ement K, Viguerie N, Poitou C, Carette C, Pelloux V, Curat CA, Sicard A, Rome S, Benis A, Zucker JD,
Vidal H, Laville M, Barsh GS, Basdevant A, Stich V, Cancello R, Langin D. Weight loss regulates
inflammation-related genes in white adipose tissue of obese subjects. FASEB J. 2004;18(14):
1657–1669.
15. Rosen ED, Spiegelman BM. What we talk about when we talk about fat. Cell. 2014;156(1-2):20–44.
16. Capel F, Klimc ´akov´a E, Viguerie N, Roussel B, V
´
ıtkov´a M, Kov ´acikov´a M, Pol ´ak J, Kov ´acov ´a Z, Galitzky
J, Maoret JJ, Han ´acek J, Pers TH, Bouloumi´e A, Stich V, Langin D. Macrophages and adipocytes in
human obesity: adipose tissue gene expression and insulin sensitivity during calorie restriction and
weight stabilization. Diabetes. 2009;58(7):1558
–1567.
17. Kov´a ˇcikov´a M, Sengenes C, Kov´a ˇcov ´aZ,
ˇ
Siklov´a-V
´
ıtkov´a M, Klim ˇc ´akov ´a E, Pol´ak J, Rossmeislov´aL,
Bajzov´a M, Hejnov ´aJ,Hnˇevkovsk ´a Z, Bouloumi´e A, Langin D,
ˇ
Stich V. Dietary intervention-induced
weight loss decreases macrophage content in adipose tissue of obese women. Int J Obes. 2011;35(1):
91–98.
18. Willett WC, Sampson L, Stampfer MJ, Rosner B, Bain C, Witschi J, Hennekens CH, Speizer FE.
Reproducibility and validity of a semiquantitative food frequency questionnaire. Am J Epidemiol. 1985;
122(1):51–65.
19. Pendyala S, Neff LM, Su´arez-Fari
~
nas M, Holt PR. Diet-induced weight loss reduces colorectal in-
flammation: implications for colorectal carcinogenesis. Am J Clin Nutr. 2011;93(2):234–242.
20. Dai Q, Gao YT, Shu XO, Yang G, Milne G, Cai Q, Wen W, Rothman N, Cai H, Li H, Xiang Y, Chow WH,
Zheng W. Oxidative stress, obesity, and breast cancer risk: results from the Shanghai Women’s Health
Study. J Clin Oncol. 2009;27(15):2482–2488.
21. Morris PG, Hudis CA, Giri D, Morrow M, Falcone DJ, Zhou XK, Du B, Brogi E, Crawford CB, Kopelovich
L, Subbaramaiah K, Dannenberg AJ. Inflammation and increased aromatase expression occur in the
breast tissue of obese women with breast cancer. Cancer Prev Res (Phila). 2011;4(7):1021–1029.
22. Iyengar NM, Morris PG, Zhou XK, Gucalp A, Giri D, Harbus MD, Falcone DJ, Krasne MD, Vahdat LT,
Subbaramaiah K, Morrow M, Hudis CA, Dannenberg AJ. Menopause is a determinant of breast adipose
inflammation. Cancer Prev Res (Phila). 2015;8(5):349–358.
23. Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data
with DESeq2. Genome Biol. 2014;15(12):550.
24. H¨
anzelmann S, Castelo R, Guinney J. GSVA: gene set variation analysis for microarray and RNA-seq
data. BMC Bioinformatics. 2013;14:7.
25. Esposito K, Pontillo A, Di Palo C, Giugliano G, Masella M, Marfella R, Giugliano D. Effect of weight loss
and lifestyle changes on vascular inflammatory markers in obese women: a randomized trial. JAMA.
2003;289(14):1799–1804.
26. Chen SB, Lee YC, Ser KH, Chen JC, Chen SC, Hsieh HF, Lee WJ. Serum C-reactive protein and white
blood cell count in morbidly obese surgical patients. Obes Surg. 2009;19(4):461–466.
636 | Journal of the Endocrine Society | doi: 10.1210/js.2017-00020
27. Keaney JF, Jr, Larson MG, Vasan RS, Wilson PW, Lipinska I, Corey D, Massaro JM, Sutherland P, Vita
JA, Benjamin EJ; Framingham Study. Obesity and systemic oxidative stress: clinical correlates of
oxidative stress in the Framingham Study. Arterioscler Thromb Vasc Biol. 2003;23(3):434–439.
28. Il’yasova D, Wang F, Spasojevic I, Base K, D’Agostino RB, Jr, Wagenknecht LE. Urinary F2-
isoprostanes, obesity, and weight gain in the IRAS cohort. Obesity (Silver Spring). 2012;20(9):
1915–1921.
29. Lumeng CN, Deyoung SM, Bodzin JL, Saltiel AR. Increased inflammatory properties of adipose tissue
macrophages recruited during diet-induced obesity. Diabetes. 2007;56(1):16–23.
30. Shapiro H, Pecht T, Shaco-Levy R, Harman-Boehm I, Kirshtein B, Kuperman Y, Chen A, Bl ¨uher M,
Shai I, Rudich A. Adipose tissue foam cells are present in human obesity. J Clin Endocrinol Metab.
2013;98(3):1173–1181.
31. Colegio OR, Chu NQ, Szabo AL, Chu T, Rhebergen AM, Jairam V, Cyrus N, Brokowski CE, Eisenbarth
SC, Phillips GM, Cline GW, Phillips AJ, Medzhitov R. Functional polarization of tumour-associated
macrophages by tumour-derived lactic acid. Nature. 2014;513(7519):559–563.
32. Pasarica M, Sereda OR, Redman LM, Albarado DC, Hymel DT, Roan LE, Rood JC, Burk DH, Smith SR.
Reduced adipose tissue oxygenation in human obesity: evidence for rarefaction, macrophage chemo-
taxis, and inflammation without an angiogenic response. Diabetes. 2009;58(3):718–725.
33. Youm YH, Nguyen KY, Grant RW, Goldberg EL, Bodogai M, Kim D, D’Agostino D, Planavsky N, Lupfer
C, Kanneganti TD, Kang S, Horvath TL, Fahmy TM, Crawford PA, Biragyn A, Alnemri E, Dixit VD. The
ketone metabolite b-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease.
Nat Med. 2015;21(3):263–269.
34. Rock CL, Emond JA, Flatt SW, Heath DD, Karanja N, Pakiz B, Sherwood NE, Thomson CA. Weight loss
is associated with increased serum 25-hydroxyvitamin D in overweight or obese women. Obesity (Silver
Spring). 2012;20(11):2296–2301.
35. Cantorna MT, Zhu Y, Froicu M, Wittke A. Vitamin D status, 1,25-dihydroxyvitamin D3, and the
immune system. Am J Clin Nutr. 2004; 80(6, Suppl)1717S–1720S.
36. Wilson PW, Abbott RD, Castelli WP. High density lipoprotein cholesterol and mortality. The Fra-
mingham Heart Study. Arteriosclerosis. 1988;8(6):737–741.
37. Winters SJ, Gogineni J, Karegar M, Scoggins C, Wunderlich CA, Baumgartner R, Ghooray DT. Sex
hormone-binding globulin gene expression and insulin resistance. J Clin Endocrinol Metab. 2014;
99
(12):E2780–E2788.
38. Magkos F, Fraterrigo G, Yoshino J, Luecking C, Kirbach K, Kelly SC, de Las Fuentes L, He S, Okunade
AL, Patterson BW, Klein S. Effects of Moderate and Subsequent Progressive Weight Loss on Metabolic
Function and Adipose Tissue Biology in Humans with Obesity. Cell Metab. 2016;23(4):591–601.
39. Iyengar NM, Hudis CA, Dannenberg AJ. Obesity and cancer: local and systemic mechanisms. Annu Rev
Med. 2015;66:297–309.
40. Campbell KL, Foster-Schubert KE, Alfano CM, Wang CC, Wang CY, Duggan CR, Mason C, Imayama I,
Kong A, Xiao L, Bain CE, Blackburn GL, Stanczyk FZ, McTiernan A. Reduced-calorie dietary weight
loss, exercise, and sex hormones in postmenopausal women: randomized controlled trial. J Clin Oncol.
2012;30(19):2314–2326.
41. James BR, Tomanek-Chalkley A, Askeland EJ, Kucaba T, Griffith TS, Norian LA. Diet-induced obesity
alters dendritic cell function in the presence and absence of tumor growth. J Immunol. 2012;189(3):
1311–1321.
42. Fallarino F, Grohmann U, You S, McGrath BC, Cavener DR, Vac ca C, Orab ona C, Bi anchi R,
Belladonna ML, Volpi C, Santamaria P, Fioretti MC, Puccetti P. The combined effects of tryptophan
starvation and tryptophan catabolites down-regulate T cell receptor zeta-chain and induce a regulatory
phenotype in naive T cells. J Immunol. 2006;176(11):6752–6761.
doi: 10.1210/js.2017-00020 | Journal of the Endocrine Society | 637
