
SPRINGER BRIEFS IN FOOD, HEALTH,
AND NUTRITION
123
Muhammad Riaz
Muhammad Zia-Ul-Haq
Bashar Saad
Anthocyanins and
Human Health:
Biomolecular
and therapeutic
aspects
SpringerBriefs in Food, Health, and Nutrition
Editor-in-chief
Richard W. Hartel , University of Wisconsin – Madison , USA
Associate Editors
John W. Finley , Louisiana State University , USA
David Rodriguez-Lazaro , ITACyL , Spain
Yrjö Roos , University College Cork , Ireland
David Topping , CSIRO , Australia
More information about this series at
http://www.springer.com/series/10203
Springer Briefs in Food, Health, and Nutrition present concise summaries of cutting
edge research and practical applications across a wide range of topics related to the
fi eld of food science, including its impact and relationship to health and nutrition.
Subjects include:
• Food chemistry, including analytical methods; ingredient functionality; physic-
chemical aspects; thermodynamics
• Food microbiology, including food safety; fermentation; foodborne pathogens;
detection methods
• Food process engineering, including unit operations; mass transfer; heating,
chilling and freezing; thermal and non-thermal processing, new technologies
• Food physics, including material science; rheology, chewing/mastication
• Food policy
• And applications to:
– Sensory Science
– Packaging
– Food Qualtiy
– Product Development
We are especially interested in how these areas impact or are related to health
and nutrition.
Featuring compact volumes of 50 to 125 pages, the series covers a range of con-
tent from professional to academic. Typical topics might include:
• A timely report of state-of-the art analytical techniques
• A bridge between new research results, as published in journal articles, and a
contextual literature review
• A snapshot of a hot or emerging topic
• An in-depth case study
• A presentation of core concepts that students must understand in order to make
independent contributions
Muhammad Riaz • Muhammad Zia-Ul-Haq
Bashar Saad
Anthocyanins and Human
Health: Biomolecular
and therapeutic aspects
ISSN 2197-571X ISSN 2197-5728 (electronic)
SpringerBriefs in Food, Health, and Nutrition
ISBN 978-3-319-26454-7 ISBN 978-3-319-26456-1 (eBook)
DOI 10.1007/978-3-319-26456-1
Library of Congress Control Number: 2015957964
© The Author(s) 2016
This work is subject to copyright. All rights are reserved by the Publisher, whether the whole or part of
the material is concerned, specifi cally the rights of translation, reprinting, reuse of illustrations, recitation,
broadcasting, reproduction on microfi lms or in any other physical way, and transmission or information
storage and retrieval, electronic adaptation, computer software, or by similar or dissimilar methodology
now known or hereafter developed.
The use of general descriptive names, registered names, trademarks, service marks, etc. in this publication
does not imply, even in the absence of a specifi c statement, that such names are exempt from the relevant
protective laws and regulations and therefore free for general use.
The publisher, the authors and the editors are safe to assume that the advice and information in this book
are believed to be true and accurate at the date of publication. Neither the publisher nor the authors or the
editors give a warranty, express or implied, with respect to the material contained herein or for any errors
or omissions that may have been made.
Printed on acid-free paper
This Springer imprint is published by Springer Nature
The registered company is Springer International Publishing AG Switzerland
Muhammad Riaz , Ph.D.
Shaheed Benazir Bhutto University
Sheringal , Pakistan
Bashar Saad , Ph.D.
Al-Qasemi Academic College
Baga Algharbiya, Israel
Arab American University Jenin
Jenin, Palestine
Muhammad Zia-Ul-Haq , Ph.D.
The Patent Offi ce
Karachi , Pakistan
v
Pref ace
During the last fi ve decades, epidemiological studies as well as basic and clinical
studies have consistently shown that there is a signifi cant positive relationship
between intake of herbs, fruits, and vegetables and reduced rate of chronic diseases
in humans, such as cardiovascular diseases, diabetes, common cancers, and other
degenerative diseases as well as aging. This is attributed to the fact that these prod-
ucts provide an optimal mix of basic macromolecules (carbohydrates, proteins,
lipids, and nucleic acids) as well as dietary fi ber, antioxidants, vitamins, and miner-
als. Various molecules in the diet can control the physiological functions of the
body and supporting immune responses. Immune functions are indispensable for
defending the body against attack by pathogens or cancer cells and thus play a
pivotal role in the maintenance of health. However, the immune functions are dis-
turbed by malnutrition, aging, physical and mental stress, or undesirable lifestyle.
Therefore, the uptake of diets with immune-modulating activities is considered an
effi cient way to prevent immune functions from declining and reduce the risk of
infection or cancer.
During the last years increasing consideration has been placed in plants and
foods which can contain antioxidant substances. The chemical compounds present
in plants that are related to health-promoting benefi ts considered several antioxi-
dants, as vitamins C and E, carotenoids, and fl avonoids. The chemical variety,
molecular weight, three-dimensional conformation, and biochemical and physical
properties of these fl avonoids allow them to interact with different targets in many
live organisms. Pigmented fl avonoids, mainly anthocyanins, are considered the
most important group of fl avonoids in plants having more than 600 compounds
identifi ed in nature. Anthocyanins are water-soluble compounds that provide color
to plant tissues (leaves, stems, roots, fl owers, and fruits) ranging from red, purple,
to blue according to the environmental pH and their structural composition.
Regarding the human consumption, the high intake of foods rich in anthocyanins
offers potential health benefi cial effects on various disorders associated with cancer,
aging diseases, obesity, neurological diseases, infl ammation, diabetes, as well as
bacterial infections.
vi
This book explores and introduces information available concerning the struc-
ture, composition, and abundance of anthocyanins in fruits and its bioavailability
and biological activity related to health-promoting effects. This book includes nine
chapters, embracing particularly historical aspects and present uses of traditional
Arab-Islamic herbal medicine. Chapter 1 focuses on botanical medicines or herbal
medicines. These therapies are still utilized as the primary form of medicine by
about 80 % of the world’s population. Over 80,000 species of plants are in use
throughout the world. Usually, leaves, fruits, fl owers, seeds, and roots are formu-
lated into tablets or pills, teas, extracts, tinctures, ointments, or creams. Currently,
about 25 % of the commonly used modern pharmaceutical drugs are of herbal origin
or contain at least one herbal-derived active compound. Indeed, some are extracted
from herbal crude extracts; others are chemically modifi ed to produce a pharmaceu-
tically active drug that agonists plant active molecule. The therapeutic effects of
medicinal plants are generally labeled as antidiabetic, anti-infl ammatory, laxative,
carminative, demulcent, antiseptic, or antitussive. Chapter 2 provides a general
introduction on anthocyanins including their general chemical structure, their thera-
peutic effi cacy, and their safety. Chapter 3 focuses on the chemical structures of
anthocyanins. It provides understanding of anthocyanins’ chemistry including their
occurrence in nature, e.g., plants and the recent discovered anthocyanins. Chapter 4
describes the utilization of anthocyanins as natural color in food and beverages.
Chapter 5 is subdivided into ten subsections, which describe intake, metabolism,
and secretions of anthocyanins in the human body. Chapter 6 provides knowledge
about the biosynthetic pathways through which these compounds are synthesized in
natural system. Chapters 7 – 9 describe the state-of-the-art knowledge of in vitro,
in vivo, and clinical literature regarding the effi cacy and safety of anthocyanins,
including anti-infl ammatory, antioxidant, antidiabetic, and anticancer effects and
prevention and treatment of degenerative diseases.
Sheringal, Pakistan Muhammad Riaz , Ph.D.
Karachi, Pakistan Muhammad Zia-Ul-Haq , Ph.D.
Jenin, Palestine Bashar Saad , Ph.D.
Preface
vii
Abbreviations
AC Anthocyanidins
ACN Anthocyanin
ACNs Anthocyanins
ACN-3-gly Anthocyanins-3-glycoside
AMPK Adenosine monophosphate protein kinase
AN-gluc Anthocyanins glucuronide
BHA Butylated hydroxyl anisole
BHT Butylated hydroxyl toluene
CBG Cytosolic B-glucosidase
CD Cluster of differentiation
CH Chalcone
C
max
Maximum concentration
CNS Central nervous system
COMT Catechol- O -methyltransferase
COX Cyclooxygenase
Cy Cyanidin
Cyd-3-glu Cyanidin-3-glucoside
Cyd-3-rut Cyanidin-3-rutinoside
Dp Delphinidin
EMIQ Enzymatically modifi ed isoquercitrin
FC Flavylium cation
FRAP Ferric reducing antioxidant potential
Gal Galactoside
Glu Glucoside
HDL High-density lipoprotein
ICAM Intracellular adhesion molecule
IL Interleukin
iNOS Inducible nitric acid synthase
LDL Low-density lipoprotein
LPH Lactase phlorizin hydrolase
LPS Lipopolysaccharide
viii
MAPK Mitogen-activated protein kinase
MCP Monocyte chemotactic protein
MCP-1 Monocyte chemotactic protein-1
MDA Malondialdehyde
MI Myocardial infarctions
Mv Malvidin
NO Nitric oxide
ORAC Oxygen radical absorbance capacity
PACNs Pyranoanthocyanins
PB Pseudo-base
PC Protocatechuic acid
PDE Phosphodiesterase
PEDF Pigment epithelial-derived factor
Pg Pelargonidin
Pn Peonidin
PPAR γ Peroxisome proliferator-activated receptor gamma
Pt Petunidin
QB Quinoidal base
RDI Recommended daily intake
R
max
Maximal rate of excretion
RNS Reactive nitrogen species
ROS Reactive oxygen species
RPE Retinal pigment epithelial
Rut Rutinoside
SAOC Serum antioxidant capacity
SGLT Sodium-glucose co-transporter
SREBP1c Sterol regulatory element-binding protein 1c
STZ Streptozocin
SULT Sulfotransferase
TAS Total antioxidant status
TBARS Thiobarbituric acid reactive substances
TEAC Trolox equivalent antioxidant capacity
TNF-α Tumor necrosis factor-α
TRAP Total reactive antioxidant potentials
UDP-GT UDP-glucuronosyltransferase
UV Ultraviolet
VEGF Vascular endothelial growth factor
VLA Very late antigen
WHO World Health Organization
Abbreviations
ix
Contents
1 Diet and Herbal-Derived Medicines ........................................................ 1
1.1 Introduction ........................................................................................ 1
1.2 Current Status of Food and Herbal-Based Medicine ......................... 3
1.3 Medicinal Plants from Tradition to Evidence-Based Application ..... 4
1.3.1 Nigella sativa ........................................................................ 5
1.3.2 Olea europaea ....................................................................... 5
1.3.3 Punica granatum ................................................................... 6
1.3.4 Trigonella foenumgraecum .................................................. 6
1.3.5 Salvia offi cinalis ................................................................... 6
1.3.6 Ammi visnaga ...................................................................... 7
1.3.7 Silybum marianum ............................................................... 7
1.3.8 Inula viscose ......................................................................... 7
1.3.9 Portulaca oleracea ................................................................ 7
1.3.10 Eruca sativa .......................................................................... 8
1.3.11 Cichorium intybus ................................................................ 8
1.3.12 Allium sativum, Garlic, and Onion (Allium cepa L.) .......... 8
1.3.13 Urtica dioica ......................................................................... 9
1.3.14 Melissa offi cinalis ................................................................ 9
1.3.15 Pimpinella anisum ................................................................ 9
1.3.16 Chamomilla recutita ............................................................. 10
1.3.17 Zingiber offi cinale ................................................................ 10
1.3.18 Rosmarinus offi cinalis ......................................................... 10
1.4 Administering Herbal-Based Treatment ............................................ 11
1.5 Herbal Active Compounds ................................................................. 11
1.6 Synergistic Actions of Foods and Phytomedicines ............................ 13
1.7 Therapeutic Properties of Herbal-Based Active Compounds ............ 14
1.8 Examples of Herbal Compounds and Pharmacological Properties ... 15
1.9 Conclusions ........................................................................................ 16
References ................................................................................................... 17
x
2 Introduction to Anthocyanins .................................................................. 21
2.1 Introduction ........................................................................................ 21
2.2 Chemical Structure ............................................................................. 23
2.2.1 Glycone Moiety and Acylating Acids .................................... 24
2.3 Pyranoanthocyanins ........................................................................... 28
2.4 Conclusions ........................................................................................ 28
References ................................................................................................... 31
3 Occurrence of Anthocyanins in Plants .................................................... 35
3.1 Introduction ........................................................................................ 35
3.2 Concentration of Anthocyanins in Fruits, Vegetable and Nuts .......... 37
3.2.1 Variation in Anthocyanins Content ........................................ 38
3.3 Typical New Anthocyanins Found in the Past Years ......................... 40
3.4 Conclusions ........................................................................................ 42
References ................................................................................................... 42
4 Anthocyanins as Natural Colors .............................................................. 47
4.1 Introduction ........................................................................................ 47
4.2 Use of Anthocyanin-Based Colorants ................................................ 51
4.3 Acylated Anthocyanins as Colorants for the Food Industry .............. 52
4.4 Conclusions ........................................................................................ 54
References ................................................................................................... 54
5 Anthocyanins Absorption and Metabolism ............................................ 57
5.1 Introduction ........................................................................................ 57
5.2 Daily Intake ........................................................................................ 58
5.3 Anthocyanins Absorption .................................................................. 59
5.3.1 Gastric Absorption ................................................................. 60
5.3.2 Absorption in the Small Intestine .......................................... 60
5.3.3 Pharmacokinetics ................................................................... 62
5.4 Carbohydrates Moieties Deconjugation ............................................. 64
5.5 The Infl uence of Colonic Microfl ora ................................................. 64
5.6 Metabolism in Intestinal Mucosa and Tissues ................................... 65
5.7 Tissue Distribution ............................................................................. 65
5.8 Excretion ............................................................................................ 66
5.9 Conclusions ........................................................................................ 67
References ................................................................................................... 67
6 Biosynthesis and Stability of Anthocyanins ............................................ 71
6.1 Introduction ........................................................................................ 71
6.2 Stability of Anthocyanins .................................................................. 73
6.3 Relationships Between Structure and Stability .................................. 74
6.4 Factor Affecting Stability of Anthocyanins ....................................... 75
6.4.1 pH ........................................................................................... 75
6.4.2 Co-pigmentation Effect .......................................................... 75
6.4.3 Solvent Effects ....................................................................... 77
6.4.4 Temperature ........................................................................... 78
Contents
xi
6.4.5 Concentration Effects ........................................................... 78
6.4.6 Oxygen ................................................................................. 78
6.4.7 Light ..................................................................................... 78
6.4.8 Enzymes ............................................................................... 79
6.4.9 Ascorbic Acid ...................................................................... 79
6.4.10 Sugars ................................................................................... 79
6.4.11 Sulfi tes.................................................................................. 80
6.5 Stability of Anthocyanins in Food Products ...................................... 80
6.6 Anthocyanins Degradation in Plants .................................................. 80
6.7 Drawback and Derivatives of Anthocyanins ...................................... 81
6.8 Anthocyanins Stabilization Mechanisms ........................................... 81
6.9 Conclusions ........................................................................................ 83
References ................................................................................................... 83
7 The Role of Anthocyanins in Health as Antioxidant,
in Bone Health and as Heart Protecting Agents..................................... 87
7.1 Introduction ........................................................................................ 87
7.2 Presumed Health-Promoting Effects of Anthocyanins ...................... 88
7.3 Toxicity of Anthocyanins ................................................................... 91
7.4 Reproductive and Developmental Toxicity ........................................ 92
7.5 Anthocyanins Biological and Pharmacological Activities ................. 92
7.6 Antioxidant Activity .......................................................................... 93
7.7 Protection Against Cardiovascular Diseases ...................................... 95
7.8 Anthocyanins and Bone Health ......................................................... 98
7.9 Conclusions ........................................................................................ 100
References ................................................................................................... 101
8 The Role of Anthocyanins in Obesity and Diabetes ............................... 109
8.1 Introduction ........................................................................................ 109
8.2 Anthocyanins and Obesity ................................................................. 110
8.3 Anthocyanins and Diabetes ................................................................ 111
8.3.1 Oxidative Stress ..................................................................... 112
8.3.2 β Cells .................................................................................... 113
8.3.3 Insulin Secretion .................................................................... 114
8.3.4 Insulin Resistance .................................................................. 115
8.3.5 α-Glucosidase Inhibitory Action ............................................ 115
8.3.6 Obesity and Diabetes ............................................................. 116
8.4 Improvement of Eye Vision ............................................................... 117
8.5 Conclusions ........................................................................................ 119
References ................................................................................................... 119
9 Anthocyanins Effects on Carcinogenesis, Immune System
and the Central Nervous System ............................................................. 125
9.1 Introduction ........................................................................................ 125
9.2 Anti-Infl ammatory Activity ............................................................... 125
9.3 Anthocyanins and Cancer .................................................................. 126
Contents
xii
9.4 Anthocyanins, Alzheimer Diseases and Brain Function .................... 131
9.5 Miscellaneous Activities .................................................................... 131
9.6 Pharmaceutical Products .................................................................... 133
9.7 Conclusions ........................................................................................ 133
References ................................................................................................... 134
Contents
1
© The Author(s) 2016
M. Riaz et al., Anthocyanins and Human Health, SpringerBriefs in Food,
Health, and Nutrition, DOI 10.1007/978-3-319-26456-1_1
Chapter 1
Diet and Herbal-Derived Medicines
1.1 Introduction
Health and food and their inter-relationship are one of most-debated topics by
people of all age and income groups and are one of the most sought after and refer-
enced topic in E-world. A peek at the magazine rack of nearby library or websites
of newspapers confi rms this as there are specifi c sections for healthy-eating there.
Healthy-diet is affordable and does not include the side effects and the metabolic
and physiologic burden that medication-packages impose on human body-systems.
Diets rich in diversifi ed eating pattern of plant-based foods are among the recom-
mended lifestyle modifi cations to decrease the risk of diseases. Fruits, nuts, herbs,
spices, grains and legumes and their products or by-products are an integral part of
the cultural, socio-economic and health systems of all countries due to their estab-
lished health-promoting effects and verifi ed immunity-boosting claims.
Botanical medicines, or herbal medicines herbal medicines also known as phyto-
therapies, are the ancient healthcare remedies known to mankind. Hundreds of wild
edible herbs and animal-derived preparations (e.g., milk, blood serum, urine, bones,
and feathers) are utilized by traditional healers to prepare remedies for the treat-
ment/prevention of all types of known illnesses as well as in maintaining healthy
body, soul, and spirit. These therapies are still utilized as the main form of drugs by
about 80 % of the world’s population. Over 80,000 species of herbs are utilized for
their medicinal properties throughout the world. Usually, leaves, fruit, fl owers,
seeds, and root are formulated into tablets or pills, teas, extracts, tinctures, ointments,
or creams [ 1 – 4 ].
The last three decades have witnessed a tremendous growth in utilization of
herbal-based diet and medicines as well as a signifi cant progress in studying risks
and benefi ts of these products at cellular and molecular levels. Herbs build an
increasingly important source of new drugs. Currently, about 25 % of the commonly
used modern pharmaceutical drugs are of herbal origin or contain at least one
herbal-derived active compound. Indeed, some are extracted from herbal crude
2
extracts; others are chemically modifi ed to produce a pharmaceutically active drug
that agonists plant active molecule. The therapeutic effects of medicinal plants are
generally labeled as anti-diabetic, anti-infl ammatory, laxative (induces bowel move-
ments or to loosen the stool), carminative (blocks the gas formation in the gastroin-
testinal tract or facilitates the expulsion of gas), demulcent (cover the mucous
membrane with soothing fi lm, healing pain and infl ammation of the membrane),
antiseptic (Healing from infections) or antitussive (cough suppressants). In contrast
to pharmaceutical medicines, which are often synthetic and usually consist of a
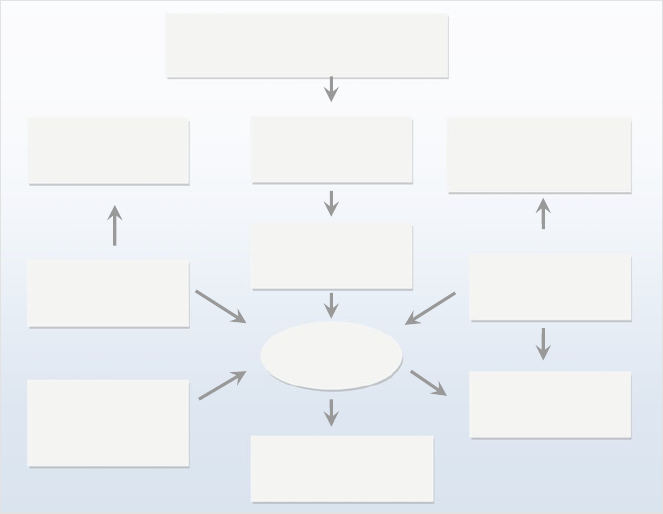
single compound), phytomedicines contain multiple constituents (Fig. 1.1 ) [ 4 – 6 ].
The popularity in utilization of the major traditional medicines has increased
worldwide over the last half century. One of the main reason for the currently
witnessed popularity is probably the belief that these medical systems have been
used for hundreds of years and that natural product-based diet and herbal-based
remedies are safe. The resurgence of interest in medicinal plant-based therapies at
the global level has been so drastic that sales of these preparations in the world are
estimated at more than 100 billion dollars per annum. Germany and France are the
principal countries in Europe in the sale of plant-based preparation. The majority of
German physicians (about 80 %) prescribe herb-based preparations [ 5 – 8 ].
Herbal medicines in the United States are sold as dietary supplements and in
many European countries are classifi ed as drugs, whereas in China and India as well
Persian, Mesopotamia
Medicine, Ancient Arabic Medicine
Innovations
introduced by Arab
and Muslim
physicians
Western medical
system
Greco-Arab
and Islamic
Persian, Chinese
Indian medicine
Pre-Islamic Arabic
medicine
Arab-Islamic medical
system
Traditional Arabic and
Islamic medicine/ Unani-
Tibb
Greekmedical system
Ayurveda and
Chinese traditional
medicines
Western traditional
medicine
Fig. 1.1 The development of various types of medical systems
1 Diet and Herbal-Derived Medicines
3
as in the Arab-Islamic world they are mostly sold over the counter without clear
regulations. One of the problems with herbal drugs is that the concentrations of
active compound(s) varies according to the soil, the weather conditions, and other
environmental factors. Safety assessments of herbal products used in these tradi-
tional medicines have often been neglected due to their prolonged and apparently
safe utilization. Nevertheless, a scientifi c evidence of the toxicity of such products
has accumulated. This is not surprising, since herbal extracts consist of mixtures of
tens of secondary metabolites, many of which are potentially toxic (e.g., hepatotox-
icity, mutagenicity, and carcinogenicity). Therefore, the extensive consumption and
fame of herbal products brought apprehensions and doubts over professionalism of
healers and quality, effi cacy, and safety of these products. Safety, contaminations,
inappropriate preparation, or lack of knowledge regarding plant and drug interac-
tions [ 6 – 9 ].
This chapter will provide a brief introduction to medicinal plants, including their
therapeutic aspects and safety. In addition, the various major types of active com-
pounds will be discussed. In the course of the following chapters, we intend to
reveal the complexities, encourage comparisons various form of anthocyanins, and
highlight relationship between their structure and functions. To keep within the
scope of this introductory chapter, we will give a brief overview of the main topics
of this book. The following chapters will comprehensively discuss the chemistry,
metabolism, in vitro and in vivo scientifi c literature as well as the clinical signifi -
cance of anthocyanins. We have organized this book around nine major topics,
refl ected by the titles of these chapters: (1) diet and herbal-derived medicines, (2)
introduction to anthocyanins, (3) occurrence of anthocyanins in plants, (4) antho-
cyanins as natural color, (5) anthocyanins absorption and metabolism, (6) biosyn-
thesis and stability of Anthocyanins, (7) the role of anthocyanins in health, (8) the
role of anthocyanins in obesity and diabetes, and (9) anthocyanins effects on carci-
nogenesis, immune system and the central nervous system.
1.2 Current Status of Food and Herbal-Based Medicine
In parallel with the revival of interest in the traditional foods and medicinal plants,
there is also an intensive research activity dealing with their safety and effi cacy as
well as with their action mechanisms at cell biological, biochemical and molecular
biological levels. The modern clinical medicine is now beginning to accept the
utilization of foods and herbal-based remedies once their effi cacy and safety are
scientifi cally investigated and validated. As a result, there is an increasing trend in
Europe as well as in the USA and Canada to incorporate herbal-derived prepara-
tions as an obligatory course in the medical curriculum. Herbalists do not isolate a
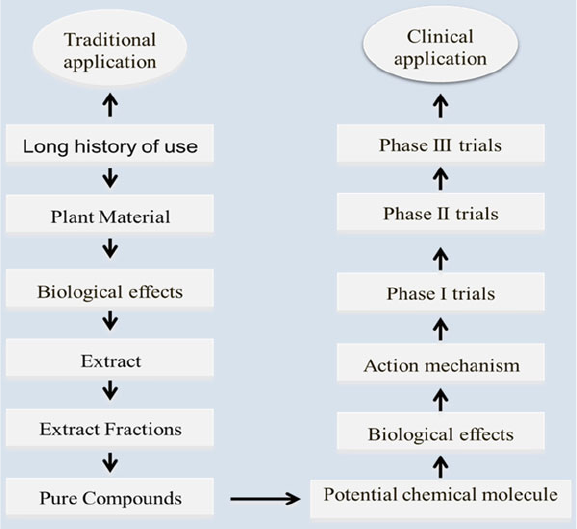
particular herbal active compound. In generally, they use the whole plant or water
extracts from parts of plants, e.g., the leaves or roots (Fig. 1.2 ). They argue that the
large number of active molecules present in herbs amplify therapeutic benefi ts and
reduce possible side effects [ 6 – 13 ].
1.2 Current Status of Food and Herbal-Based Medicine
4
Currently, products from biological sources, mainly from plants, animals, fungi,
and algae represent a main source of new medicines. The World Health Organization
(WHO) estimates that about 80 % of the world population presently utilizes plant-
based preparations for prevention or treatment of all known diseases. Although
fi nancial funding for natural product–based drug development in the pharmaceutical
industry has been reduced from 1984 to 2003, the percentage of natural product–
based substances has remained relatively at relatively high level. Approximately
70 % of anticancer drugs developed since 1940s (155 drugs) have their origin from
natural products [ 5 – 8 , 13 ].
1.3 Medicinal Plants from Tradition to
Evidence-Based Application
Medicinal plants are classifi ed as wild-grown or as cultivated. As a result of natural
selection, wild medicinal plants are grown in places with optimal soil and environ-
mental conditions. These herbs may have been exposed to environmental pollution
and pesticides. Hence, cultivated organic herbs grown under well controlled
Fig. 1.2 Commonly used preparation procedure
1 Diet and Herbal-Derived Medicines
5
conditions represent a better alternative to wild-grown herbs. Organic vegetables
and fruits are becoming increasingly available, as more and more farmers start to
adapt the organic system. With careful management, organic food farms can provide
high quality organic products to the market. In the case of medicinal plants, it is
important that medicinal plant as well as fruits and vegetables have been harvested
at the optimum season in order to achieve optimal therapeutic effects. For instance,
many edible wild plants are collected in the spring, winter, and fall, but not in
summer, when energy of plants is utilized for the purposes of fl owering and growth.
To keep within the scope of this chapter we will give a brief overview of the most
commonly used medicinal herbs in the Mediterranean region where more than 2600
plant species are found and about 450–700 plants are mentioned in various scripts
for their uses as medicinal herbs. Plant parts used include seeds, roots, fl owers,
leaves, stems and fruits [
1 – 5 , 12 – 14 ].
1.3.1 Nigella sativa
The seeds of Nigella sativa (black seeds) are commonly used medicinal herbs
throughout the Arab counties. These seeds have been used since dawn of civiliza-
tion as a spice and food preservative and for the treatment and prevention of a wide
range of diseases. Thymoquinone, dithymquinone, thymohydroquinone, and thymol
are the main bioactive constituents responsible for the pharmacological properties
of black seeds [
14 , 15 ]. Pharmacological and toxicological effects of the seeds have
been extensively studied. In recent years, scientifi c journals have published
numerous articles pointing to potential therapeutic properties Nigella sativa ,
including antimicrobial, hypotensive, antidiabetic, anticancer, antihistaminic,
immunomodulatory and anti-fertility effects [ 8 , 14 – 17 ].
1.3.2 Olea europaea
The olive, like Nigella sativa , is one of the most commonly utilized medicinal herbs
throughout the Mediterranean. Both, olive oil and olive leaf are well known for their
health benefi ts and have been for these properties for thousands of years. The
primary medical constituents of olive leaf are the antioxidants oleuropein, hydroxy-
tyrosol, hydroxytyrosol acetate, and the fl avonoids luteolin, and luteolin- glucosides.
Oleuropein and its hydrolysis products are those of the greatest therapeutic potential.
Oleuropein has a vasodilator effect, increases blood fl ow in the coronary arteries
and improves arrhythmia. It has proven to be a potent anti- infl ammatory and
antioxidant compound. Various scientifi c reports show that oleuropein also exhibits
in antimicrobial activity against viruses, retroviruses, bacteria, yeasts, fungus,
molds and other parasites [
8 , 14 , 18 , 19 ].
1.3 Medicinal Plants from Tradition to Evidence-Based Application
6
1.3.3 Punica granatum
The pomegranate has been used since long in Arab and Islamic traditional medicine
to treat a wide range of diseases. These include anticancer, anti-infl ammatory and
anti-rheumatic activities. In Ayurvedic medicine the fruits as well as other parts of
the plant are regarded as “remedy for every disease”. For example, the bark and
roots are used to manage vermifuge and helminthic conditions and the juice is
considered as “refrigerant” and “blood tonic”. Pomegranate fl owers serve as a
remedy for diabetes mellitus. The hydrolyzable tannins known as punicalagins
show anti- oxidant effects in in vitro experiments. A Medline search using therapeu-
tic potential and toxicological effects Punica granatum discloses greater than 350
citations, including antioxidant, cardiovascular protection, oral hygiene, as well as
anti- infl ammatory properties [ 8 , 14 , 20 ].
1.3.4 Trigonella foenumgraecum
Locally known as fenugreek, it is one of the most used herb in the Mediterranean.
Most of the medicinal benefi ts of Trigonella foenum - graecum are found in the
seeds, which have been used for thousands of years in Chinese and Ayurvedic
medical systems. The anti-diabetic effects of fenugreek are mediated through the
active compound, 4-hydroxy isoleucine. The action mechanisms include
translocation of glucose transporter-4 (GLUT4) to the plasma membrane, delay of
gastric emptying, slowing glucose absorption, and transport from the fi ber content,
as well as increased erythrocyte insulin receptors and modulation of peripheral
glucose utilization [ 8 , 14 , 21 , 22 ].
1.3.5 Salvia offi cinalis
Commonly known as sage, pharmacological and toxicological properties of Salvia
offi cinalis leaf have been extensively investigated using in vitro and in vivo tests. The
main active compounds of sage are present in its essential oil, which contains thu-
jone, borneol, and cineole. In addition, sage leaf contains ursonic acid, tannic acid,
ursolic acid, oleic acid, chlorogenic acid, fumaric acid, cornsolic acid, caffeic acid,
niacin, nicotinamide, fl avonoid glycosides, and estrogenic compounds. Salvia offi ci-
nalis exhibits a wide spectrum of pharmacological effects. These includes antioxidant,
anti-infl ammatory, antimicrobial, carminative, weakly spasmolytic, astringent, and
antihidrotic (inhibits perspiration). Salvia offi cinalis is also a well-known tonic and
stimulant for the nervous and digestive systems. Clinical studies have demonstrated
benefi cial therapeutic properties of Salvia offi cinalis leaves in elderly patients
suffering from mild to moderate Alzheimer’s disease [ 8 , 14 , 23 – 25 ].
1 Diet and Herbal-Derived Medicines
7
1.3.6 Ammi visnaga
Khella is used to treat breathing system disorders such as asthma, bronchitis, whoop-
ing cough, cardiac diseases and liver and gall bladder disorders and is believed to help
discharge of kidney stones and gallstones. Khella contains coumarins and furocouma-
rins the most signifi cant being khellin, visnadin and visnagin. Visnadin exhibits
coronary and peripheral vasodilatory activities in isolated vascular smooth muscle of
rat model. The coronary, urological and respiratory clinical and therapeutic effective-
ness of khellin is well-established. Furthermore, khella has been found effective in the
treatment of mild angina complaints, postoperative treatment of urinary calculus and
supportive treatment of mild forms of obstructive pulmonary diseases [ 8 , 14 , 26 – 28 ].
1.3.7 Silybum marianum
Milk thistle is used in liver supporting functions, treatment of chronic and acute
liver disease, as well as promoting detoxifying functions of the liver. The active
compounds of Silybum marianum are fl avonolignans including silidianin, silybin
and silichristine, jointly recognized as silymarin. Silybin is the molecule with the
greatest degree of pharmacological effects. Silymarin protects against several types
of xenobiotic including alcohol. Medical studies have verifi ed the pharmacological
effects of standardized milk thistle extracts in cases of cirrhosis, toxic liver and
allied liver conditions [ 8 , 14 , 29 ].
1.3.8 Inula viscose
Tayun, has been used traditionally as one of the most effective medicinal herb for
the treatment of infections, infl ammations, various skin diseases and wound healing.
The roots have been utilized to treat cough, phlegm and sepsis. The leaves contain
essential oils, fl avonoids such as rhamnocitrin, glycosyl analogue of diacylglycerol,
sakuranetin, methylaromadendrin, acetyl-methylaromadendrin, and sesquiterpene
lactones. Inula viscosais well documented to have anti-ulcerogenic effects and to
cause abortion in mammals. Further effects include antimicrobial and fungi effects,
anti-infl ammatory and anti-diabetic properties. Due to these wide range of medical
effects, this plant is appreciated by pharmaceutical industries [ 8 , 30 – 33 ].
1.3.9 Portulaca oleracea
Purslane, is used traditionally in the treatment of a wide range illnesses, such as
include headache, stomach ache, enteritis, painful urination, mastitis. It is also used
to increase milk production in nursing women and in treating postpartum bleeding.
1.3 Medicinal Plants from Tradition to Evidence-Based Application
8
Arial parts of this plan have therapeutic effects in healing burns, earache, ulcers, and
pruritis. They are also used to treat infl ammations, skin sores, insect stings, eczema,
and wound healing. Purslane contains relatively high levels of a neurohormone
( L -norepinephrine) that was found to exhibit vasopressor and anti-hypotensive and
anti-haemorrhage effects. Various in vitro and in vivo studies indicate that aqueous
extracts of this plant exhibit skeletal muscle relaxant effects [ 3 , 4 , 8 ].
1.3.10 Eruca sativa
Rucola, is considered traditionally as a general tonic and potent aphrodisiac.
Additional traditional uses of Eruca sativa include stimulation of spermatogenesis
and fertility, antibacterial effects, and promoting kidney function and digestion.
Eruca sativa extract was found to exhibit signifi cant antioxidant properties.
Glucoerucin and fl avonoids are the major antioxidants present in Eruca sativa .
Feeding of Eruca sativa extract to rats induced a signifi cant protection against HgCl
induced renal toxicity. In addition, there are several scientifi c reports that indicate
that Eruca sativa exhibit antimicrobial effect [ 8 , 14 , 34 , 35 ].
1.3.11 Cichorium intybus
Wild chicory, fi nds a widespread use both in the inhibition and treatment of a wide
spectrum of illnesses. It is useful in promoting liver functions (detoxifying functions)
as well as encouraging the eliminative pathways both through the intestine and the
kidneys. Arabic traditional healers recommend chicory as part of a combined treat-
ment of metabolic problems, colds, and fl u. The roots of chicory contain inulin and
oligofructose polysaccharides. Chicory, like many plants that support liver function
and immunity, has strong antioxidant effects in vitro, but the clinical signifi cance of
this has not been tested. However there have been several studies in humans on the
therapeutic effects of the inulin and oligofructan polysaccharides. They have been
found to undergo fermentation in the colon and to selectively stimulate of the growth
of healthy bifi dobacteria population that results in the decrease of colonic diseases
and diabetes, as well as support for the immune system [ 14 , 36 , 37 ].
1.3.12 Allium sativum , Garlic, and Onion ( Allium cepa L.)
These are one of the most used plants for their well-known health benefi ts. Garlic
has been used for centuries for prevention and treatment of a large number of
illnesses. Chinese as well as Greeks and Romans utilized onion and garlic-based
preparations for the treatment and prevention of diseases. The two plants are rich
1 Diet and Herbal-Derived Medicines
9
sources of large number of active compounds. Onion and garlic is important ingre-
dient of the Mediterranean diet. Various scientifi c reports indicate their effi cacy in
the treatment and prevention of a wide range of pathological conditions, such as
cancer, cardiovascular diseases and obesity [ 3 , 4 , 8 , 14 ].
1.3.13 Urtica dioica
Stinging nettle is highly appreciated in Greco-Arab medicine for its benefi cial
effects. These include anti-rheumatic effects, anti-colds and anti-cough, and
promoting liver functions, anti-hypotensive and anti-infl ammatory effects.
Evidence- based therapeutic application of this plant includes anti-diabetes antioxi-
dant, anti-infl ammatory, increasing cell growth lymphocytes in humans, anti-
prostatic hyperplasia and anti-hypertension [ 8 , 14 , 38 , 39 ].
1.3.14 Melissa offi cinalis
Lemon balm, the therapeutic uses of this plant dates back into ancient times. Greco-
Arab and Islamic physicians used the herb to treat heart disorders. Melissa offi cina-
lis leaves contain about 0.1 % of essential oil, consisting of a highly variable mixture
of constituents. These include and polyphenolic compounds (mainly rosmarinic
acid and monoterpene glycosides), monoterpenoid aldehydes, and fl avonoids.
Currently, Melissa offi cinalis is highly appreciated by traditional healers. The herb
fi nds a widespread use in the treatment of skin diseases (mainly acne). Additional
therapeutic effects include sedative effect on the central nervous system of mice,
antimicrobial and antiviral effects, anti-hyperthyroidism and anti-depression effects.
Lemon palm has also positive effects on the nervous system. In addition to anti-
depression effects on patients, the plant is effective in decreasing symptoms of
Alzheimer’s and dementia such as memory loss [ 14 ].
1.3.15 Pimpinella anisum
Anise, as cumin, fennel, carrots, cilantro, and dill belong to the Apiaceae family.
The seeds (“fruits”) are used traditionally to treat of a wide range of illnesses,
particularly for their benefi cial effects in reduction of problems related to digestion.
Seed-based therapies are commonly used with babies and children to heal from
baby colic. Furthermore, these seeds are also recommended by traditional healers to
treat symptoms associated with indigestion and nausea. In additional, their antispas-
modic effect is one of most known therapeutic property of anise. The seeds
commonly used to treat menstrual pain, asthma attacks, whooping cough and other
1.3 Medicinal Plants from Tradition to Evidence-Based Application
10
spasmodic coughs. Furthermore, anise seed-based preparations are rationally used
for their ability to increase the production of milk in nursing mothers. Anise-derived
essential have the same therapeutical properties as the whole seeds. Women in the
fi rst term of pregnancy must should avoid taking anise [ 8 , 14 ].
1.3.16 Chamomilla recutita
Chamomile is appreciated for the medicinal benefi ts of the essential oils and
infusions prepared from fl ower heads. Due to their aromatic, fl avoring and coloring
properties they fi nd a widespread use in commercial products including liniments,
balms, hair products, soaps, detergents, perfumes, bakery and confectionary
products, and herbal teas. Phenolic compounds, primarily the fl avonoids quercetin,
patuletin, apigenin and luteolin are the main active compounds of the fl owers.
Medicinal benefi ts of Chamomile-based remedies include antioxidant, antimicro-
bial activities and antiplatelet activity. Animal model investigations verify
anti-infl ammatory effects, anti-mutagenic and cholesterol-lowering effects, as well
as anti-spasmotic and anxiolytic properties [ 8 , 14 ].
1.3.17 Zingiber offi cinale
The rhizome (the underground stem) of the ginger is appreciated globally both as
spice and for its medicinal properties to treat arthritis, colic, diarrhea, painful men-
strual pains, as well as common cold and fl u. The rhizomes have been utilized since
ancient times as a one of the effective herbal remedies in various systems of medi-
cine. Currently, traditional healers recommend ginger for the treatment/prevention
of nausea and vomiting related with pregnancy, cancer and motion sickness. In
addition, rhizome-based extracts fi nd a wide spread use as a gastrointestinal utility
for minor stomach troubles and to cure infl ammations like arthritis [ 8 , 14 ].
1.3.18 Rosmarinus offi cinalis
Rosemary, the areal parts of this highly aromatic plant, known for its bitter and
astringent taste, are customarily utilized all over the Mediterranean region, both as
cooking spice and for their medicinal properties. Rosemary contains a number of
bioactive phytochemicals, such as the antioxidants carnosic acid and rosmarinic
acid. Rosemary is known for its effects on muscle relaxation. Because of this prop-
erty it is conventionally used to relieve digestive problems and to ease menstrual
pains. A tea made from the leaves is also taken as a tonic for calming nerves and
used as an antiseptic. Several studies showed that carnosic acid, found in rosemary,
exhibit a strong antioxidant and antimicrobial properties [
14 , 40 ].
1 Diet and Herbal-Derived Medicines

11
1.4 Administering Herbal-Based Treatment
Several preparation methods were developed in major traditional medicines are still
practiced by traditional herbalists to prepare herbal-based medicines. The majority
of herbal preparations are used as tea or water diluted extracts. Heating fresh or
dried plant parts in a solvent result in the extraction of bioactive phytochemical. In
addition, this procedure helps to reduce or even to eliminate impurities and poisons
and prior to application (Table 1.1 ). The chemical composition and concentration of
an extract is largely affected by the solvent used in the extraction. Water extracts
will be rich in hydrophilic phytochemicals, oil on the other hand will absorb hydro-
phobic substances. Alcohol will help in extracting, both polar and un-polar
compounds. Other extraction methods include the inhalation of aerosols, essential
oils (Essential oils are volatile, complex, natural compounds formed by aromatic
plants), tinctures (tinctures are preparations containing alcohol), capsules and
tablets and vaporized plant juices or teas [ 1 – 5 , 41 ].
1.5 Herbal Active Compounds
Plants produce metabolites as part of their normal cellular metabolic functions.
These are classifi ed as primary metabolites, present in all plants, and secondary
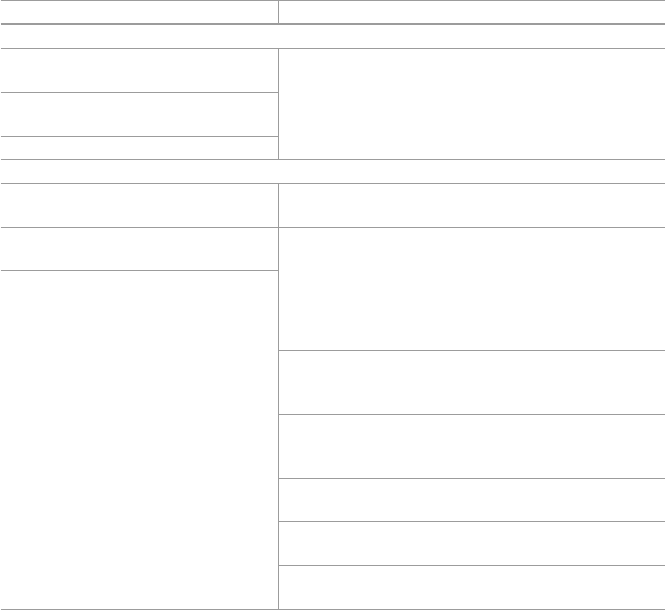
metabolites eliciting pharmacological effects in man and animals (Table 1.2 ). Basic
metabolism comprises all primary metabolites essential for the survival of the plant
which are involved in the primary anabolic and catabolic cellular processes respon-
sible for types of cellular activities (e.g., cell growth and differentiation). In con-
trast, secondary metabolites are those that found usually only in special, differentiated
cells/tissues and are not necessary for the cells/tissue themselves but are important
for the plant as a whole. The number of known secondary metabolites that have
been discovered to date is increasing at a constant rate. Yet, it is not only plants that
produce these bioactive compounds; rather, other organisms such as fungi, bacteria,
Table 1.1 Preparations methods used for oral administrations
Administration form Preparation methods
Whole plant Fresh juice; fresh/dried areal parts and other underground parts
Tinctures Preparations of plant extract with varying ratios of water and alcohol
Tisanes Hot water extracts of plants
Decoctions and teas Made by steeping and soaking herb (leaves, fl owers, stems, roots,
and bark) in water for a few minutes
Vinegars Prepared as tinctures
Syrups Extracts of herbs made with syrup or honey
Extracts Extracts are liquids with a lower alcohol level than tinctures
Essential oils Essential oil extracts are usually diluted in carrier oil
1.5 Herbal Active Compounds
12
sponges, as well as animals, are also capable of synthesizing a large number of these
compounds. In general, secondary metabolites often possess interesting therapeutic
properties in humans and animals, and therefore their investigation is very important.
It should not be forgotten that plants synthesize these compounds as part of their
own survival strategies. For example, some secondary products are pheromones
used to attract insects for pollination, while others are toxins used to deter predation.
Phytoalexins protect the plant against fungal or bacterial infections. Flavonoids acts
as antioxidants to neutralize free radicals generated during photosynthesis.
Anthocyanins may attract pollinators or seed dispersers. Alkaloids can protects
against herbivore animals or insect attacks. Plants regulate their cellular metabolism
in response to the present herbivores, pollinators, microorganisms, and other
environmental stresses. In addition, recent evidence has pointed to additional roles
for secondary metabolites in plant development. Although the term “ secondary
metabolites” perhaps infers a less important role for these compounds than those
involved in primary metabolism, this is not the case. In fact, many essential and
nonessential compounds in this group are found in plants, and even so-called
“ nonessential materials” can play a role in a plant’s responses against abiotic and
biotic stress.
Table 1.2 Secondary metabolites and their properties
Metabolites Examples
Primary metabolites :
Organic compounds produced in
plants
Polypeptides, cellulose, amino acids, nucleic acids,
mono-saccharides, and lipids
Essential for basic cell growth and
differentiation
Produced all plant tissues
Secondary metabolites :
Organic compounds produced in
plants
Generally grouped into classes:
Do not have essential role involved
in growth and differentiation
Polyphenols (Widely distributed in the plant
kingdom, responsible for the colors of many
fl owers, others are present in bark, roots and leaves
that play an important role in tanning hides and
skins to give leather. Yet others are simpler
compounds found in most fresh fruit and vegetables
Produced in different plant
families, in specifi c groups of plant
families or in specifi c tissues, cell/
tissue specifi c, produce at different
developmental stages and in
response to environmental stresses
Terpenoids and steroids are derived
biosynthetically from isopentenyldiphosphate).
Over 35,000 compounds are known
Fatty acid - derived substances and polyketides are
biosynthesized from simple acyl precursors such as
acetyl CoA. More than 10,000 molecules are known
Alkaloids are derived biosynthetically from amino
acids. More than 12,000 compounds are known
Nonribosomal polypeptides are biosynthesized
from amino acids
Enzyme cofactors are coenzymes such as pyridoxal
phosphate
1 Diet and Herbal-Derived Medicines
13
In general, secondary metabolites occur as complex mixtures. The chemical com-
position and concentration of same plant can vary over time in response to variation
in environmental conditions. Their biosynthesis can also be infl uenced by a variety
of factors during development, in addition to stress, which makes the determination
of their complete pattern essentially very diffi cult. Whilst secondary metabolites can
occur in the tissues as active compounds, they can also be synthesized as inactive
compounds that must be transformed into active products. Compounds that are bio-
synthesized under stress conditions are typically not detectable in unstressed tissues;
when they are synthesized after the invasion of plants by various pests. The patterns
of secondary metabolites will differ depending on the species. The synthesis of
secondary metabolites can occur in all plant organs, including the roots, shoots,
leaves, fl owers, fruit, and seeds. Some metabolites are stored in specifi c compart-
ments, which may be either whole organs or specialized cell types. Within these
compartments the concentration of toxic secondary metabolites may be very high, so
that they can exert an effi cient defense against herbivores.
In order to identify or quantify a compound of interest, the metabolite must fi rst
be extracted from the plant tissues. However, the chemical properties of a material
under investigation isof great importance in the development of a relevant purifi ca-
tion scheme (Fig. 1.1 ). The most important issues to be taken into account include:
It must be defi ned whether a compound or a broad range of already known should
be extracted and quantifi ed. In addition, for individual compounds, it must be
determined which properties are already known, and which solvents can be used for
their extraction. And fi nally, the purity of the compound might be important for
identifi cation and also for bioactivity assays; in this situation the metabolite must be
further purifi ed using chromatographic methods [ 5 , 7 , 40 , 42 ].
1.6 Synergistic Actions of Foods and Phytomedicines
In contrast to synthetic drugs based upon one pure active molecule, the majority of
herbal-derived medicines exert their pharmacological action via synergistic or
additive pathway of a mixture active biomolecules acting at single or multiple target
tissues associated with a pathophysiological pathway. In addition to desired thera-
peutical action these synergistic and additive effects can be advantageous by
reducing negative side effects allied with the use of drugs consisting of a single
pharmaceutical molecule. Additive and synergistic effects likely have their origin in
the physiological and metabolic roles of secondary products in stimulating plant
survival, regeneration and growth. For instance, a combination of secondary metab-
olites having additive or synergistic action at multiple target cell/tissue would not
only guarantee effi cacy in fi ghting wide range of pathogens and herbivores but
would also reduce or even eliminate the probabilities of these pathogens developing
adaptive responses or resistance [ 11 – 14 ].
1.6 Synergistic Actions of Foods and Phytomedicines
14
1.7 Therapeutic Properties of Herbal-Based Active
Compounds
Plants synthesize a wide range of secondary metabolites but most are derived from
a few chemical motifs. These phytochemicals can have pharmacologic properties in
humans and can be chemically modifi ed to produce new medicines. Numerous
herbal-derived substances have been investigated for their pharmacologic potential
as new drugs. These include fl avonoids, coumarins, saponins and alkaloids.
Flavonoids, in particular anthrocyanins, are probably the best elucidated phyto-
chemicals of these biomolecules due to their potent antioxidant activity. The medical
benefi t of numerous plant herbal-based remedies used by traditional healers, at least
in part, is attributed to their effective antioxidant effects.
As above discussed, Black seed has been used for centuries in Greco-Arab and
Islamic medicine for its magic healing properties as well as its disease prevention
effects. Avicenna (980–1037 AC) highlighted the medical benefi ts of black seeds
that they act as energy-booster of human body and serves to recover from fatigue
and dispiritedness. Thymoquinone presents the main active molecule responsible
for the biological and pharmacological properties of black seed. It was found to
inhibit a wide range of pathogenic processes. For example, antioxidant, immun-
emodulatory, anti-cancer, hypolipidemic, and vasoconstrictive properties in cell
culture systems and animal models. Additional therapeutical properties of black
seeds include, inhibition of iron-dependent microsomal lipid peroxidation, cardio-
toxicity induced by doxorubin in rats, drug-induced toxicity and ameliorates the
anticancer effects. One of the very important pharmacological properties of thymo-
quinone is its high cytotoxic effects as assessed in canine osteosarcoma, colon
cancer, skin cancer and prostate cancer. In contrast thymoquinone showed low
cytotoxicity to normal cells. Thymoquinone also cures many multidrug-resistant
types of pancreatic adenocarcinoma, human leukemia and uterine sarcoma.
Furthermore, many in vitro and in vivo mechanistic studies indicate that thymoqui-
none induces apoptosis through affecting multiple cellular and biochemical targets.
Therefore, this compound present a promising example of phytochemical that is
helpful for the prevention and treatment of many types of cancer cells. This antican-
cer property was also supported by studies in prostate and other cancer cells.
Thymoquinone was found to inhibit angiogenesis in vivo, prohibited tumor angio-
genesis in a xenograft human prostate cancer model in mouse and blocked human
prostate tumor growth without any side effects. Thymoquinone also exhibits
anti- proliferative activity in colon and prostate tumors implanted in nude mice.
Taken together, these fi nding show that the anticancer and cytostatic properties are
due to the effect of thymoquinone on cell cycle [ 18 , 19 ]. In addition, these results
indicate a great potential for the development of new synthetic derivatives of thymo-
quinone as anticancer drugs [ 43 – 45 ].
Another example of potential group phytochemical is anthocyanins. This phyto-
chemicals are one of the most abundant fl avonoid compounds and one of the most
widespread families of natural pigments in the plant kingdom. These pigments,
1 Diet and Herbal-Derived Medicines
15
present in fruits and vegetables, provide color and promote health benefi ts to con-
sumers due to their antioxidant capacity. To date, more than 600 anthocyanins have
been identifi ed in the plant kingdom. The different anthocyanin absorb light at about
500 nm and are responsible for the red, blue and purple color of fruits and vegeta-
bles. All known anthocyanins conjugates are based on six anthocyanidin aglycones
derived from fl avylium backbone with different glycosylations and acylations. As
discussed in the coming chapters of this book, many studies in cell lines, animal
models and human clinical trials suggest that anthocyanins have anti-carcinogenic
and anti-infl ammatory activities, provides cardiovascular disease prevention,
promote obesity and diabetes control benefi ts, and also improve visual and brain
functions. Those health benefi ts are mainly associated with their antioxidant effects,
which clearly are infl uenced by the molecular mechanism related to the expression
and modulation of key genes.
1.8 Examples of Herbal Compounds and Pharmacological
Properties
Many drugs currently listed as conventional medications are prepared from herbal-
derived active compounds. The majority of these herbal-derived medicines were
discovered by the study of the old traditional medical systems, namely, the Chinese,
Ayurvadic, and Greco-Arab medicine. For instance, the cardiac glycoside obtained
from the foxglove ( Digitalis purpurea ) are the most cited molecules of herbal-
derived drugs for treatments of cardiovascular diseases. They are matchless by any
synthetic or semi-synthetic medicines and have exceptional effi cacy with selective
cardiotonic activity. Another example is the study of cardiovascular properties of
herbs that led to the discovery of reserpine over 65 years ago. Reserpine is derived
from the roots of Rauwolfi a serpentine and Vakil in 1949 reported it as a hyperten-
sive drug. About 10 years later, reserpine was isolated said plant, its structure was
elucidated and it was synthesized in labs. Later on, reserpine was found to be a
potent agent in treating Parkinson disease and depression. These results stimulated
further scientifi c research and it was observed that reserpine decreased brain
nor- epinephrine, dopamine as well as serotonin. This was a break-through in
research on transmitter amine defects in depression and Parkinson’s disease. This
was a milestone for the development of many psychoactive drugs and led to a
substantial interaction between scientists and pharmacological industry.
Other examples of herbs as a source of pharmaceutical active compounds
include: Vincristine is obtained from Periwinkle and used as an anti-cancer remedy.
Cinchona bark is the source of malaria-fi ghting quinine. For centuries, herbalists
prescribed echinacea obtained from purple conefl ower to fi ght infection. This herb
was one of the most extensively recommended medicines in the US before the
discovery and synthesis of antibiotics. Now it has been confi rmed that echinacea
improves the immune system by increasing the generation of lymphocytes. Willow
bark-derived salicylic acid (Aspirin) is a key anti-infl ammatory, antipyretic and
1.8 Examples of Herbal Compounds and Pharmacological Properties
16
analgesic molecule frequently used in clinical medicine. Another example of herbal-
derived medicines is opium poppy ( Papaver somniferum )-derived morphine which
is one of the early compounds used in conventional medicine systems and is a
premium painkiller. The isolation of morphine from crude opium by Serturner in
1806 stirred so much research on herabal drugs that Megendie published a medical
formulary in 1821, containing only pure chemical entities, hence laid paving the
pathway for the use of single and pure compounds instead of medicinal plants and
their extracts [ 43 – 45 ].
1.9 Conclusions
In parallel with the increasing in utilization of nutraceuticals and herbal-based
medicines, there is also an intensive scientifi c research activity dealing with their
safety and health benefi cial effects. These include management of wide range of
diseases as well as in elucidation of their action mechanisms in vitro, in vivo and
in clinical studies. Plants metabolites are classifi ed as primary metabolites, like
proteins and lipids which are found in all plants, and secondary metabolites elicit-
ing pharmacological effects in man and animals. Basic metabolism comprises all
primary metabolites necessary for the survival of the cells, which are found in all
plants and are responsible for the primary metabolic functions of building and
sustaining plant cells. In contrast, secondary plant metabolites are those that occur
usually only in special, differentiated tissues and are not necessary for the cells
themselves but are important for the plant as a whole. The number of known sec-
ondary metabolites that have been discovered to date is increasing at a constant
rate. Herbal-derived medicines exercise their pharmacological actions through the
synergistic or additive pathway of numerous active molecules that act at single or
many target tissue linked with a physiological pathway. Many drugs currently
listed as conventional medications are prepared from herbal-derived active com-
pounds. These herbal-derived drugs were discovered through the study of tradi-
tional medical systems, namely, the Chinese, Ayurvadic, and Greco-Arab medicine.
Anthocyanins are one of the most abundant fl avonoid compounds and one of the
most widespread families of natural pigments in the plant kingdom. The following
chapters will comprehensively discuss the chemistry, metabolism, in vitro and
in vivo scientifi c literature as well as the clinical signifi cance of anthocyanins.
Regarding pharmacological properties of anthocyanins, a lot needs to be eluci-
dated. Understand their action mechanisms in the prevention of chronic diseases,
cancer, neurodegenerative diseases, and aging are still to be unveiled. As we will
see in following chapters of this book, investigations regarding absorption and
distribution anthocyanins are still needed. Furthermore, the effect of long-term
exposure to anthocyanins is still largely uninvestigated and more in depth in vivo
and clinical studies are needed in order to elucidate implications of anthocyanins
in the before mentioned health-promoting effects.
1 Diet and Herbal-Derived Medicines
17
References
1. Saad, B., Azaizeh, H., & Said, O. (2008). Arab herbal medicine. Botanical Medicine in
Clinical Practice, 4 , 31.
2. Saad, B., Azaizeh, H., & Said, O. (2005). Tradition and perspectives of Arab herbal medicine:
A review. Evidence-Based Complementary and Alternative Medicine, 2 (4), 475–479.
3. Saad, B., & Said, O. (2011). Herbal medicine. In B. Saad & O. Said (Eds.), Greco-Arab and
Islamic herbal medicine: Traditional system, ethics, safety, effi cacy and regulatory issues
(pp. 47–71). Hoboken: Wiley.
4. Saad, B. (2014). Greco-Arab and Islamic herbal medicines, a review. European Journal of
Medicinal Plants, 4 (3), 249–258.
5. Si-Yuan, P., Shu-Feng, Z., Si-Hua, G., Zhi-Ling, Y., Shuo-Feng, Z., Min-Ke, T., et al. (2013).
New perspectives on how to discover drugs from herbal medicines: CAM’s outstanding con-
tribution to modern therapeutics. Evidence-Based Complementary and Alternative Medicine .
doi:
10.1155/2013/627375 .
6. Costa-Neto, E. M. (2005). Animal-based medicines: Biological prospection and the sustain-
able use of zootherapeutic resources. Anais da Academia Brasileira de Ciências, 77 (1), 33–43.
7. Li, J. W.-H., & Vederas, J. C. (2009). Drug discovery and natural products: End of an era or an
endless frontier? Science, 325 (5937), 161–165.
8. Saad, B. (2015). Integrating traditional Greco‐Arab and Islamic diet and herbal medicines in
research and clinical practice. In I. Ramzan (Ed.), Phytotherapies: Effi cacy, safety, and regula-
tion (p. 142). Hoboken: Wiley.
9. Saad, B., Azaizeh, H., Abu-Hijleh, G., & Said, O. (2006). Safety of traditional Arab herbal
medicine. Evidence-Based Complementary and Alternative Medicine, 3 (4), 433–439.
10. Pormann, P. E., Savage-Smith, E., & Hehmeyer, I. (2007). Medieval Islamic medicine .
Edinburgh: Edinburgh University Press.
11. Cragg, G. M., & Newman, D. J. (2005). Biodiversity: A continuing source of novel drug leads.
Pure and Applied Chemistry, 77 (1), 7–24.
12. Saad, B., & Said, O. (2011). The current state of knowledge of Arab herbal medicine. In
B. Saad & O. Said (Eds.), Greco-Arab and Islamic herbal medicine: Traditional system, ethics,
safety, effi cacy, and regulatory issues . Hoboken: Wiley.
13. Harvey, A. L. (2008). Natural products in drug discovery. Drug Discovery Today, 13 (19),
894–901.
14. Saad, B., & Said, O. (2011). Commonly used herbal medicines in the mediterranean. In
B. Saad & O. Said (Eds.), Greco-Arab and Islamic herbal medicine: Traditional system, ethics,
safety, effi cacy, and regulatory issues (pp. 149–227). Hoboken: Wiley.
15. Salem, M. L., & Hossain, M. S. (2000). Protective effect of black seed oil from Nigella sativa
against murine cytomegalovirus infection. International Journal of Immunopharmacology,
22 (9), 729–740.
16. Gilani, A., Jabeen, Q., & Khan, M. A. U. (2004). A review of medicinal uses and pharmaco-
logical activities of Nigella sativa. Pakistan Journal of Biological Sciences, 7 , 441–451.
17. Ghosheh, O. A., Houdi, A. A., & Crooks, P. A. (1999). High performance liquid chromato-
graphic analysis of the pharmacologically active quinones and related compounds in the oil of
the black seed ( Nigella sativa L.). Journal of Pharmaceutical and Biomedical Analysis, 19 (5),
757–762.
18. Omar, S. H. (2008). Olive: Native of Mediterranean region and health benefi ts. Pharmacognosy
Reviews, 2 (3), 135–142.
19. Yaseen Khan, M., Siddharth, P., Niraj, V., Amee, B., & Vimal, K. (2007). Olea europaea : A
phyto-pharmacological review. Pharmacognosy Reviews, 1 (1), 114–118.
20. Lansky, E. P., & Newman, R. A. (2007). Punica granatum (pomegranate) and its potential for
prevention and treatment of infl ammation and cancer. Journal of Ethnopharmacology, 109 (2),
177–206.
21. Raju, J., Gupta, D., Rao, A. R., Yadava, P. K., & Baquer, N. Z. (2001). Trigonella foenum
graecum (fenugreek) seed powder improves glucose homeostasis in alloxan diabetic rat tissues
References
18
by reversing the altered glycolytic, gluconeogenic and lipogenic enzymes. Molecular and
Cellular Biochemistry, 224 (1–2), 45–51.
22. Kadan, S., Saad, B., Sasson, Y., & Zaid, H. (2013). In vitro evaluations of cytotoxicity of eight
antidiabetic medicinal plants and their effect on GLUT4 translocation. Evidence-Based
Complementary and Alternative Medicine, 2013 , 549345.
23. Hohmann, J., Zupkó, I., Rédei, D., Csányi, M., Falkay, G., Máthé, I., et al. (1999). Protective
effects of the aerial parts of Salvia offi cinalis , Melissa offi cinalis and Lavandula angustifolia
and their constituents against enzyme-dependent and enzyme-independent lipid peroxidation.
Planta Medica, 65 (6), 576–578.
24. Zupkó, I., Hohmann, J., Rédei, D., Falkay, G., Janicsák, G., & Máthé, I. (2001). Antioxidant
activity of leaves of Salvia species in enzyme-dependent and enzyme-independent systems of
lipid peroxidation and their phenolic constituents. Planta Medica, 67 (4), 366–368.
25. Kennedy, D. O., Pace, S., Haskell, C., Okello, E. J., Milne, A., & Scholey, A. B. (2006). Effects
of cholinesterase inhibiting sage ( Salvia offi cinalis ) on mood, anxiety and performance on a
psychological stressor battery. Neuropsychopharmacology, 31 (4), 845–852.
26. Rauwald, H. W., Brehm, O., & Odenthal, K.-P. (1994). The involvement of a Ca2+ channel
blocking mode of action in the pharmacology of Ammi visnaga fruits. Planta Medica, 60 (2),
101–105.
27. Carlie, G., Ntusi, N. B., Hulley, P. A., & Kidson, S. H. (2003). KUVA (khellin plus ultraviolet
A) stimulates proliferation and melanogenesis in normal human melanocytes and melanoma
cells in vitro. British Journal of Dermatology, 149 (4), 707–717.
28. Hofer, A., Kerl, H., & Wolf, P. (2001). Long-term results in the treatment of vitiligo with oral
khellin plus UVA. European Journal of Dermatology, 11 (3), 225–229.
29. Ball, K. R., & Kowdley, K. V. (2005). A review of Silybum marianum (milk thistle) as a treat-
ment for alcoholic liver disease. Journal of Clinical Gastroenterology, 39 (6), 520–528.
30. Máñez, S., Recio, M. C., Gil, I., Gómez, C., Giner, R. M., Waterman, P. G., et al. (1999). A
glycosyl analogue of diacylglycerol and other antiinfl ammatory constituents from Inula vis-
cosa. Journal of Natural Products, 62 (4), 601–604.
31. Ali-Shtayeh, M. S., Yaghmour, R. M., Faidi, Y. R., Salem, K., & Al-Nuri, M. A. (1998).
Antimicrobial activity of 20 plants used in folkloric medicine in the Palestinian area. Journal
of Ethnopharmacology, 60 (3), 265–271.
32. Maoz, M., Kashman, Y., & Neeman, I. (1999). Isolation and identifi cation of a new antifungal
sesquiterpene lactone from Inula viscosa. Planta Medica, 65 (3), 281–282.
33. Tripathi, Y., Tripathi, P., & Upadhyay, B. (1988). Assessment of the adrenergic beta-blocking
activity of Inula racemosa. Journal of Ethnopharmacology, 23 (1), 3–9.
34. Jirovetz, L., Smith, D., & Buchbauer, G. (2002). Aroma compound analysis of Eruca sativa
( Brassicaceae ) SPME headspace leaf samples using GC, GC-MS, and olfactometry. Journal
of Agricultural and Food Chemistry, 50 (16), 4643–4646.
35. Lamy, E., Schröder, J., Paulus, S., Brenk, P., Stahl, T., & Mersch-Sundermann, V. (2008).
Antigenotoxic properties of Eruca sativa (rocket plant), erucin and erysolin in human hepa-
toma (HepG2) cells towards benzo (a) pyrene and their mode of action. Food and Chemical
Toxicology, 46 (7), 2415–2421.
36. Kisiel, W., & Zielińska, K. (2001). Guaianolides from Cichorium intybus and structure revi-
sion of Cichorium sesquiterpene lactones. Phytochemistry, 57 (4), 523–527.
37. Roberfroid, M. B. (1999). Concepts in functional foods: The case of inulin and oligofructose.
The Journal of Nutrition, 129 (7), 1398S–1401S.
38. Gülçin, I., Küfrevioglu, O. I., Oktay, M., & Büyükokuroglu, M. E. (2004). Antioxidant, anti-
microbial, antiulcer and analgesic activities of nettle ( Urtica dioica L.). Journal of
Ethnopharmacology, 90 (2), 205–215.
39. Farzami, B., Ahmadvand, D., Vardasbi, S., Majin, F. J., & Khaghani, S. (2003). Induction of
insulin secretion by a component of Urtica dioica leave extract in perifused Islets of Langerhans
and its in vivo effects in normal and streptozotocin diabetic rats. Journal of Ethnopharmacology,
89 (1), 47–53.
1 Diet and Herbal-Derived Medicines
19
40. Salem, M. L. (2005). Immunomodulatory and therapeutic properties of the Nigella sativa L.
seed. International Immunopharmacology, 5 (13), 1749–1770.
41. Bakkali, F., Averbeck, S., Averbeck, D., & Idaomar, M. (2008). Biological effects of essential
oils – A review. Food and Chemical Toxicology, 46 (2), 446–475.
42. Gali-Muhtasib, H., Diab-Assaf, M., Boltze, C., Al-Hmaira, J., Hartig, R., Roessner, A., et al.
(2004). Thymoquinone extracted from black seed triggers apoptotic cell death in human
colorectal cancer cells via a p53-dependent mechanism. International Journal of Oncology,
25 (4), 857–866.
43. Ji, H. F., Li, X. J., & Zhang, H. Y. (2009). Natural products and drug discovery. EMBO Reports,
10 (3), 194–200.
44. Amin, A., Gali-Muhtasib, H., Ocker, M., & Schneider-Stock, R. (2009). Overview of major
classes of plant-derived anticancer drugs. International Journal of Biomedical Science, 5 (1),
1–10.
45. Briskin, D. P. (2000). Medicinal plants and phytomedicines. Linking plant biochemistry and
physiology to human health. Plant Physiology, 124 (2), 507–514.
References
21
© The Author(s) 2016
M. Riaz et al., Anthocyanins and Human Health, SpringerBriefs in Food,
Health, and Nutrition, DOI 10.1007/978-3-319-26456-1_2
Chapter 2
Introduction to Anthocyanins
2.1 Introduction
Medicinal plants and diet therapies are the oldest methods of prevention and treat-
ment of all types of known diseases and built a substantial part of the main tradi-
tional medical systems in maintaining healthy body, soul, and spirit. The past three
decades have witnessed signifi cant increase in utilization as well as a big progress
in studying the cellular and molecular interactions between intake of healthy foods
(e.g., edible wild plants, nuts, seeds, fruits, vegetables as well as olive oil) and
reduced rate of diabetes, common cancers, cardiovascular diseases, ageing and
degenerative diseases. The terms functional foods and nutraceuticals refer to phy-
tochemical substances that have long-term health benefi cial effects. Although there
is very thin difference between medicinal plants and functional foods, the latter also
known as super-foods provide health-promoting and immunity boosting effects
besides providing optimum nutrition with minimum amounts. The use of medicinal
plants however leads to specifi c biological actions without any nutritional aspect.
These pharmacological effects of medicinal plants, their extracts or their formula-
tions are due to secondary metabolites also known as bioactive compounds or phy-
tochemicals present in them. These secondary metabolites are specifi c for plant
species and families and also serve a role in taxonomic identity of their respective
species and families. The secondary metabolites differ from primary metabolites
such as lipids, proteins, carbohydrates and nucleic acids which are commonly
found in all plants and are responsible for primary metabolism and cell growth and
regeneration. Various plant secondary metabolites, also known as active compounds
can regulate the metabolism and physiological functions of the human body, and
support immune responses. As discussed in Chaps. 7 , 8 , and 9 , both arms of the
immune system, namely the innate and the adaptive systems are indispensable for
defending the body against attack by pathogens or cancer cells, and thus play a
pivotal role in the maintenance of health. Hence, the intake of herbs and foods with
22
immunity-modulating effects are an effective way to inhibit deterioration of immu-
nity potential and decrease the chances of allied disorders like cancer, infl ammation
or infection [ 1 – 3 ].
The term anthocyanin is coined from the Greek anthos , meaning fl ower, and
kyanos , meaning blue in 1835 by a German scientist Ludwig Marquart as previ-
ously they were known as colored cell sap, cyanophylls, chrythophylls or cyanins.
Earlier it was believed that these colors arise from the degradation of chlorophyll
during autumn. Boyle in 1664 in his research paper entitled, “Experiments and
considerations touching colors” for the fi rst time noticed that color change of the
“syrup of violet” from intense purple to green is due to “acid liquor”. The fi rst book
ever written was, “The anthocyanin pigments of plants” by Muriel Wheldale in
1916. Willstatter and Everest (1913), Willstatter and Nolan (1915) and Shibata
et al. (1919) performed fi rst time experiments on pH dependent changes of antho-
cyanin colors of Centaurea cyanus fl owers. This group reported for the fi rst time
that development of C. cyanus petal color is due to conjugation of anthocyanins
with metal ions [ 4 – 9 ].
The molecular structure of anthocyanins was discovered by Richard Willstatter
and his colleagues from 1912 to 1916 shortly after the rediscovery of Mendel’s
laws of inheritance. In this era, anthocyanins were subject of molecular genetics
since Mendel’s pea had distinct color due to anthocyanins [ 10 ]. The color dis-
played by these constituents was described by Pauling for the fi rst time in 1939
who suggested that the resonate structure of the fl avylium cation causes intensity
of their color. These are the biggest group of vacuolar natural colors that are water
soluble [ 11 ] and are the most important pigments followed by chlorophylls that
are visible to human eye. Their color is due to their resonating structure which is
also the basis of their instability. Being phytochemicals, these molecules cannot
be synthesized by humans and animals and they have to rely on plants for their
requirements. Anthocyanins are found in many plant tissues predominantly in the
epidermal tissues, palisade, spongy mesophyll of leaves, fl esh of fruits/stem and
underground storage organs [ 12 ]. but most commonly accumulated in fl owers and
fruits [ 13 ]. Anthocyanins provide color to plant tissues (fl owers, stems, leaves,
roots) ranging from red, purple to blue according to the pH and their structural
composition.
The anthocyanins are recently the focus of substantial basic and pharmacological
research because of their sparkling color, high water solubility and valuable biologi-
cal properties. They are less potent as compared to corresponding medications, but
since they are consumed in substantial amounts in regular diet, they exert clearly
visible long-term biological effects. The French paradox revealed that French popu-
lation has less chances of coronary heart disease because of more consumption of
red wine. The plethora of available fl ora in region and their possible combinations
used in making this wine provides a diverse and rich pool of anthocyanins. These
compounds play an important role in human in prevention and treatment of a wide
range of diseases. Now-a-days, anthocyanins are becoming an integral part of human
diet. Nutritionists and diet-counselors are increasingly recommending the use of
2 Introduction to Anthocyanins
23
anthocyanin-rich foods as well as pure anthocyanins to treat many diseases, due to
their proven ability to simulate specifi c hormones and neurotransmitters, to inhibit
some enzymes, and to act as antioxidant. They also act as a secondary antioxidant
defense system in plant tissues exposed to different abiotic and biotic environmental
stresses like fungal pathogens, UV light, cold temperature and dry weather [ 14 ].
2.2 Chemical Structure
The understanding of the chemistry of anthocyanins is of vital importance in assess-
ment their biological and pharmacological effects. Although hundreds of anthocya-
nins have been identifi ed, only a small fraction of these molecules has been studied
in depth. Each anthocyanin has its specifi c and precise three dimensional structures
which ensures optimal molecular fi tting into specifi c cell and sub-cellular binding
sites leading to biological effects. The biological properties of anthocyanins depend
on their chemical structure, substitutions, conjugations and polymerization. In
nature, these exist as glycosides of fl avylium salts and 90 % of identifi ed anthocya-
nins are based on six major anthocyanidins (aglycone). They differ from each other
depending upon sugar moiety attached to the fl avyliumn cation and to the acylation,
hydroxylation and methoxilation pattern. Spectroscopic, molecular and functional
genomic studies have exhibited remarkable similarity among all anthocyanins in
sense as they all share a common skeleton. Basic skeleton structure of anthocyanins
is shown in Fig. 2.1 .
Chemically, anthocyanins belong to the fl avonoid group that are glycosylated
polyhydroxy and polymethoxy derivatives (3,5,7,39-tetrahydroxyfl avylium cation)
of fl avilium salts, possessing a characteristic C-6 (A ring)-C-3 (C ring)-C-6 (B ring)
carbon structure [ 14 ]. Structurally, these are heterosides of an aglycone unit (antho-
cyanidin) which is a derivative of the fl avylium ion. The general formula of antho-
cyanins is
O
R
4
R
3
R
2
R
1
R
7
R
6
R
5
1
2
3
4
5
6
7
8
1`
2`
3`
4`
5`
6
`
A
C
B
Fig. 2.1 Basic skeleton
structure of anthocyanins
2.2 Chemical Structure
24
Anthocyanidins sugars Anthocyanins+Þ
The aglycone (anthocyanidins) part of the anthocyanins are quite reactive due
to electron defi cient fl avylium cation and has lower solubility so they occur
always as glycosides in nature i.e. bonded to glycosyl moiety to make it more
stable than the aglycones. The most common glycone moiety are D -glucose,
L -rhamnose, D - galactose, D -xylose and arabinose, these moieties are usually
located at carbons 3, 5, 7, 3′, and 5′ [ 15 , 16 ]. Glycosylation at C-3 position is
most common as compared to other position [ 17 ]. All anthocyanins are type
O -glycosides i.e. the sugar substituent is attached through O linkage [ 15 ]. Various
factors affect their stability in food including processing and storage temperature,
pH, content and identity in food matrix, oxygen, enzymes and metallic ions.
Anthocyanins are different from each other by number of hydroxylated groups in
the anthocyanidin, the nature and the number of bonded sugars in their structure,
the aliphatic or aromatic carboxylates bonded to the sugar in the molecule, and
the position of these bonds [ 18 ]. The variation of methoxylation and hydroxyl-
ation patterns in these structures produces hues from orange-red (pelargonidin) to
blue-violet (delphinidin) at pH ≈ 1. Remarkably, the anthocaynins behave in same
way in vivo : nasturtium fl owers ( Tropaeolum majus L.), radish ( Raphanus sativus
L.) and strawberry ( Fragaria ananassa Duch.) are orange-red due to pelargonidin
derivatives, while larkspur petals ( Delphinium consolida L.), blueberries
( Vaccinium sp.) and grapes ( Vitis sp.) are blue-violet due to delphinidin-type
anthocyanins [ 19 ]. Due to their water solubility, they are incorporated into aque-
ous food systems. Anthocyanins occurring in nature contain several anthocyani-
dins or aglycones, but only six are common in foods e.g. cyanidin, peonidin,
pelargonidin, malvidin, delphinidin, and petunidin [ 20 ]. The structures of some
common anthocyanins are given in Fig. 2.2 .
2.2.1 Glycone Moiety and Acylating Acids
Glucose, arabinose, rhamnose, galactose, xylose or glucuronic acid are the reported
monosaccharaides chains that are linked to C-3/5/7/3′/4′/5′, which are often acyl-
ated by aliphatic acids such as acetic, malonic, malic, oxalic, tartaric and succinic
acids and also by cyclic acids like caffeic, sinapic, ferulic, gallic, p-hydroxybenzoic
and p-coumaric acids, concurrently up to three acylating acids may be present [ 21 ].
Pendant sugars increase stabilization, and absorption of these small molecules
(±500 g/mol) into human cells and their small size helps in penetrating the blood-
brain barrier and cell walls. Some common monosaccharides that are linked to agly-
cone moiety are shown in Fig. 2.3 while structures of some acylated cyclic and
aliphatic acids are depicted in Figs. 2.4 and 2.5 , respectively.
2 Introduction to Anthocyanins
25
Fig. 2.2 Various anthocyanins isolated from natural sources
O
+
H
O
H
OH
H
H
O
H
H
Apigeninidin (Ap)
O
+
H
O
H
O
H
H
O
O
H
H
Arrabidin (Ab)
O
+
H
O
HO
O
H
OH
H
O
H
H
Aurantinidin (Au)
O
+
H
O
H
O
O
H
O
O
H
O
Capensinidin (Cp)
O
+
H
O
HO
H
H
O
O
H
Carajurin (Cj)
O
+
H
O
H
OH
OH
H
O
H
O
H
Cyanidin (Cy)
O
+
H
O
H
OH
OH
OH
O
H
O
H
Delphinidin (Dp)
O
+
H
O
H
O
OH
OH
OH
O
O
+
O
H
OH
OH
O
OH
O
Europinidin (Eu)
Hirsutidin (Hs)
O
+
H
O
H
OH
H
H
OH
O
H
Luteolin (Lt)
O
+
H
O
H
OH
OH
O
O
H
O
Malvidin (Mv)
O
+
H
O
H
O
OH
H
OH
OH
5-MethylCyanidin (5-MCY)
O
+
H
O
H
OH
O
H
H
O
H
H
Pelargonidin (Pg)
O
+
H
O
H
OH
O
H
H
O
H
O
Peonidin (Pn)
O
+
H
O
H
OH
O
H
O
H
O
H
O
Petunidin (Pt)
(continued)
2.2 Chemical Structure
O
H
O
OH
O
H
OH
H
O
T
T
H
O
O
OH
O
H
OH
G
lucose Ar
a
binose
O
O
H
OH OH
O
H
Rhamnose
O
H
O
O
H
O
H
OH
OH
Galactose
HO OH
H
O
O
OH
Xylose
O
H
OH
OH
HO
O
H
O
O
Glucuronic acid
Fig. 2.3 Monosaccharides
that are linked to aglycone
O
+
H
O
H
O
OH
OH
O
H
O
H
Pulchellidin (Pl)
O
+
H
O
HO
H
OH
H
O
H
H
Riccionidin A (RiA)
O
+
O
H
OH
OH
H
OH
O
Rosinidin (Rs)
O
+
H
O
H
OH
H
OH
O
H
O
H
Tricetinidin (Tr)
O
+
H
O
H
O
H
H
O
O
H
O
H
30-HydroxyAb (3'OHAb)
O
+
H
O
H
O
OH
OH
OH
O
H
O
H
6-HydroxyCy (6OHCy)
O
+
H
O
HO
OH
OH
OH
OH
O
H
6-HydroxyDp (6OHDp)
O
+
H
O
HO
OH
OH
H
OH
H
6-HydroxyPg (6OHPg)
Fig. 2.2 (continued)
27
Fig. 2.4 Acylated aliphatic acids
O
OH
OH
HO
Ca
ffi
e
i
cac
id
HO
O
O
OH
O
S
i
na
pi
cac
i
d
O
HO
O
OH
F
e
r
u
li
c
a
c
i
d
O
OH
OH
H
O
HO
Gallic acid
OH
O
HO
p-hydroxybenzoic acid
OH
O
HO
p-coumaric acid
Fig. 2.5 Acylated cyclic acids
2.2 Chemical Structure
28
2.3 Pyranoanthocyanins
Pyranoanthocyanins are the products of anthocyanins and low molecular weight
such as pyruvic acid, 4-vinylphenol and fl avonols. They are stable derivatives of
anthocyanins and considered to contribute the age-related color changes in plants,
and the fi rst pyranoanthocyanins was reported in 1996 in red wines fi ltrates [ 22 ].
Some pyranoanthocyanins are also detected in black carrot juice and in blood
orange juices [ 22 , 23 ]. Pyranoanthocyanins are stable anthocyanins derivatives that
were identifi ed in red grape wines [ 24 ]. The cyclo-addition of anthocyanins and
ethylenic bond of another molecule at C-4 and C-5 results in the formation of bleach
and pH change resistant, pyranoanthocyanins [ 25 , 26 ]. Rosacyanin B violet was the
fi rst pyranoanthocyanidin isolated from plant petals Rosa hybrida .
These derivatives of anthocyanins have important qualities in terms of stability
to gastro environment may results in more pharmacological active molecule and
also results more stable color to change of temperature, pH or other chemical reac-
tion like bleaching with SO
2
. Figure 2.6 indicates structure of various reported
pyranoanthocyanins.
For example, Vistins, a group of pyranoanthocyanins in wine with orange-red
color, exhibit 11 and 14 times greater color at wine pH, than malvidin-3- O - glucoside
[ 27 ]. Almost all reported pyranoanthocyanins have orange color except portisins
which has blue color. Lu et al. (2002) isolated four pyranoanthocyanins, namely
pyranodelphinin C and D and pyranocyanin C and D, possessing the pyrano[4,3,2-
de ]-1-benzopyrylium core structure from an extract of blackcurrant seeds [ 28 ].
Table 2.1 shows various reported anthocyanins.
Pyranoanthocyanins have also been observed in small quantity in extracts of red
onions [ 21 ] and strawberries [ 38 ]. Four reported methyl pyranoanthocyanins iso-
lated from black currant seeds were identifi ed as the oxidative cyclo-addition prod-
ucts of the anthocyanins and extraction solvent i.e. acetone [ 39 ]. It is confi rmed
from studies that pyaranoanthocyanins are more stable than their parent anthocya-
nins, so a new strategy would be to increase the stability of anthocyanins, the impor-
tance of this group has already been discussed in previous chapters.
2.4 Conclusions
Most living matter in nature has color and a major percentage of these colors are
plant-derived. Anthocyanins are incorporated in food chains because of their pres-
ence in fruits and vegetables and their corresponding beverage products like juices,
jams, jellies, wines, confectionary and bakery products as food color, functional
food or as a dietary supplement. Anthocyanins have aromatic rings containing polar
substituent groups (–OH, –OCH
3
) and glycosyl residues making them a polar mol-
ecule. Structurally these are benzopyran derivatives; i.e. oxygenated heterocyclic
compounds while chemically these are chromophores having conjugated double
2 Introduction to Anthocyanins

29
Fig. 2.6 Structure of various reported pyranoanthocyanins
O
O
COOH
H
O
OGlu
R
1
OH
R
2
Vitisins A
O
O
HO
OGlu
R
1
O
H
R
2
Vitisins B
O
O
H
O
OGlu
R
1
O
H
R
2
Methyl pyranoanthocyanins
O
O
HO
OGlu
R
1
OH
R
2
O
10-acetylpyranoanthocyanins
O
O
H
O
OGlu
R
1
O
H
R
2
O
O
HO
OGlu
R
1
OH
R
2
R
3
R
4
O
H
O
HO
OH
O
H
OH
OH
Pinotins
Flavanylpyranoanthcyanins
(continued)
2.4 Conclusions
30
O
O
H
O
OGlu
R
1
O
H
R
2
O
H
O
OH
OH
OH
OH
Portisins A
O
O
H
O
OGlu
R
1
O
H
R
2
R
4
OH
Portisins B
R
3
O
O
HO
OGlu
R
1
O
H
R
2
O
Oxovitisins
O
O
H
O
O
G
l
u
O
O
H
O
OO
OH
OGlu
O
H
O
O
Pyranoanthocyanin dimer
O
O
OH
OH
H
O
O
OH
O
O
O
OH
OH
OH
OH
Pinotin A
Fig. 2.6 (continued)
2 Introduction to Anthocyanins
31
bonds. Due to the technological advancements in instruments and chemical deter-
minations, considerable information has been gained on the identifi cation and char-
acterization of anthocyanins. Despite so much progress, still hundreds of
anthocyanins remain unidentifi ed and so full potential of many plant species has not
been explored yet. More work is required to isolate and characterize remaining
anthocyanins and to decipher their biological activities as it will provide invaluable
insights into mechanism underlying their health-promoting and immunity boosting
effects.
References
1. Saad, B., & Said, O. (Eds.). (2011). Greco-Arab and Islamic herbal medicine: Traditional
system, ethics, safety, effi cacy, and regulatory issues . Hoboken: Wiley.
2. Saad, B. (2015). Integrating traditional Greco‐Arab and Islamic diet and herbal medicines in
research and clinical practice. In I. Ramzan (Ed.), Phytotherapies: Effi cacy, safety, and regula-
tion (p. 142). Hoboken: Wiley.
3. Saad, B. (2015). Greco-Arab and Islamic diet therapy: Tradition, research and practice.
Arabian Journal of Medicinal and Aromatic Plants, 1 , 1–24.
4. Boyle, R. (2004). Experiments and considerations touching colours (1664) . London: Henry
Herringman.
5. Wheldale, M. (1916). Anthocyanic pigments of plants . Cambridge: Cambridge University
Press.
6. Willstätter, R., & Everest, A. E. (1913). Untersuchungen über die Anthocyane. I. Über den
Farbstoff der Kornblume. Justus Liebigs Annalen der Chemie, 401 (2), 189–232.
7. Willstätter, R., & Nolan, T. J. (1915). Untersuchungen über die Anthocyane. II. Über den
Farbstoff der Rose. Justus Liebigs Annalen der Chemie, 408 (1), 1–14.
Table 2.1 Various types of reported pyranoanthocyanins
Cyclo-addition Type of pyaranoanthocyanin References
Anthocyanin + pyruvic acid Carboxypyranoanthocyanins (Vitisin A) [
29 ]
Anthocyanin + acetaldehyde Pyranoanthocyanins (Vitisin B) [
30 ]
Anthocyanins + pyruvic
acid + fl avanols in the presence of
acetaldehyde
Portisins [
31 ]
Anthocyanins + pyruvic
acid + vinylphenol + phloroglucinol
Portisins [
32 ]
Anthocyanins + pyruvic
acid + phenol
Portisins, oxovitisins, and
pyranoanthocyanin dimmers
[
33 ]
Anthocyanins + pyruvic
acid + acetone
Methyl-pyranoanthocyanin [
34 ]
Anthocyanins + diacetyl 10-acetylpyranoanthocyanins [
35 ]
Anthocyanins + p -coumaric or
sinapic acids
Pinotins
(hydroxyphenylpyranoanthocyanins)
[
36 ]
Pyranoanthocyanin + fl avanols Flavanylpyranoanthocyanins [
32 ]
Anthocyanins + 4-vinylcatechol Pinotin A [
37 ]
References
32
8. Shibata, K., Shibata, Y., & Kasiwagi, I. (1919). Studies on anthocyanins: Color variation in
anthocyanins. Journal of the American Chemical Society, 41 (2), 208–220.
9. Landi, M., Tattini, M., & Gould, K. S. (2015). Multiple functional roles of anthocyanins in
plant-environment interactions. Environmental and Experimental Botany, 119 , 4–17.
10. Lee, D., & Gould, K. (2002). Why leaves turn red pigments called anthocyanins probably
protect leaves from light damage by direct shielding and by scavenging free radicals. American
Scientist, 90 (6), 524–528.
11. Ghosh, D., & Konishi, T. (2007). Anthocyanins and anthocyanin-rich extracts: Role in diabetes
and eye function. Asia Pacifi c Journal of Clinical Nutrition, 16 (2), 200.
12. Oren-Shamir, M. (2009). Does anthocyanin degradation play a signifi cant role in determining
pigment concentration in plants? Plant Science, 177 (4), 310–316.
13. Delgado-Vargas, F., & Paredes-López, O. (2003). Anthocyanins and betalains. Natural
Colorants for Food and Nutraceutical 167–219.
14. Chalker Scott, L. (1999). Environmental signifi cance of anthocyanins in plant stress responses.
Photochemistry and Photobiology, 70 (1), 1–9.
15. Da Costa, C. T., Horton, D., & Margolis, S. A. (2000). Analysis of anthocyanins in foods by
liquid chromatography, liquid chromatography-mass spectrometry and capillary electrophore-
sis. Journal of Chromatography A, 881 (1), 403–410.
16. Strack, D., Steglich, W., & Wray, V. (1993). Betalains. Methods in Plant Biochemistry, 8 ,
421–450.
17. Bkowska-Barczak, A. (2005). Acylated anthocyanins as stable, natural food colorants – A
review. Polish Journal of Food and Nutrition Sciences, 14 , 107–116.
18. Valls, J., Millán, S., Martí, M. P., Borràs, E., & Arola, L. (2009). Advanced separation methods
of food anthocyanins, isofl avones and fl avanols. Journal of Chromatography A, 1216 (43),
7143–7172.
19. Stintzing, F. C., & Carle, R. (2004). Functional properties of anthocyanins and betalains in
plants, food, and in human nutrition. Trends in Food Science and Technology, 15 (1), 19–38.
20. Kong, J. M., Chia, L. S., Goh, N. K., Chia, T. F., & Brouillard, R. (2003). Analysis and biologi-
cal activities of anthocyanins. Phytochemistry, 64 (5), 923–933.
21. Andersen, Ø. M., Fossen, T., Torskangerpoll, K., Fossen, A., & Hauge, U. (2004). Anthocyanin
from strawberry ( Fragaria ananassa ) with the novel aglycone, 5-carboxypyranopelargonidin.
Phytochemistry, 65 (4), 405–410.
22. Rentzsch, M., Schwarz, M., & Winterhalter, P. (2007). Pyranoanthocyanins – An overview on
structures, occurrence, and pathways of formation. Trends in Food Science and Technology,
18 (10), 526–534.
23. Castañeda-Ovando, A., Pacheco-Hernández, M., Páez-Hernández, M. E., Rodríguez, J. A., &
Galán-Vidal, C. (2009). Chemical studies of anthocyanins: A review. Food Chemistry, 113 (4),
859–871.
24. Pozo-Bayón, M. A., Monagas, M., Polo, M. C., & Gómez-Cordovés, C. (2004). Occurrence of
pyranoanthocyanins in sparkling wines manufactured with red grape varieties. Journal of
Agricultural and Food Chemistry, 52 (5), 1300–1306.
25. Monagas, M., Bartolomé, B., & Gómez-Cordovés, C. (2005). Evolution of polyphenols in red
wines from Vitis vinifera L. during aging in the bottle. European Food Research and
Technology, 220 (3–4), 331–340.
26. Bakker, J., & Timberlake, C. F. (1997). Isolation, identifi cation, and characterization of new
color-stable anthocyanins occurring in some red wines. Journal of Agricultural and Food
Chemistry, 45 (1), 35–43.
27. Romero, C., & Bakker, J. (1999). Interactions between grape anthocyanins and pyruvic acid,
with effect of pH and acid concentration on anthocyanin composition and color in model solu-
tions. Journal of Agricultural and Food Chemistry, 47 (8), 3130–3139.
28. Lu, Y., Foo, L. Y., & Sun, Y. (2002). New pyranoanthocyanins from blackcurrant seeds.
Tetrahedron Letters, 43 (41), 7341–7344.
29. Fulcrand, H., Montserrat, D., Erika, S., & Véronique, C. (2006). Phenolic reactions during
winemaking and aging. American Journal of Enology and Viticulture, 57 (3), 289–297.
2 Introduction to Anthocyanins
33
30. Bakker, J., & Timberlake, C. F. (1985). The distribution of anthocyanins in grape skin extracts
of port wine cultivars as determined by high performance liquid chromatography. Journal of
the Science of Food and Agriculture, 36 (12), 1315–1324.
31. Mateus, N., Oliveira, J., Haettich-Motta, M., & de Freitas, V. (2004). New family of bluish
pyranoanthocyanins. BioMed Research International, 2004 (5), 299–305.
32. de Freitas, V., & Mateus, N. (2011). Formation of pyranoanthocyanins in red wines: A new and
diverse class of anthocyanin derivatives. Analytical and Bioanalytical Chemistry, 401 (5),
1463–1473.
33. Schwarz, M., Wabnitz, T. C., & Winterhalter, P. (2003). Pathway leading to the formation of
anthocyanin-vinylphenol adducts and related pigments in red wines. Journal of Agricultural
and Food Chemistry, 51 (12), 3682–3687.
34. Mateus, N., Pascual-Teresa, S., Rivas-Gonzalo, J. C., Santos-Buelga, C., & Victor de Freitas,
V. D. (2002). Structural diversity of anthocyanin-derived pigments in port wines. Food
Chemistry, 76 (3), 335–342.
35. Gómez-Alonso, S., Collins, V. J., Vauzour, D., Rodríguez-Mateos, A., Corona, G., & Spencer,
J. P. E. (2012). Inhibition of colon adenocarcinoma cell proliferation by fl avonols is linked to
a G2/M cell cycle block and reduction in cyclin D1 expression. Food Chemistry, 130 (3),
493–500.
36. He, F., Liang, N. N., Mu, L., Pan, Q. H., Wang, J., & Reeves, M. J. (2012). Anthocyanins and
their variation in red wines I. Monomeric anthocyanins and their color expression. Molecules,
17 (2), 1571–1601.
37. Schwarz, M., Jerz, G., & Winterhalter, P. (2015). Isolation and structure of Pinotin A, a new
anthocyanin derivative from Pinotage wine. VITIS-Journal of Grapevine Research, 42 (2), 105.
38. Fossen, T., & Andersen, Ø. M. (2003). Anthocyanins from red onion, ( Allium cepa ) with novel
aglycone. Phytochemistry, 62 (8), 1217–1220.
39. Lu, Y., & Foo, L. Y. (2001). Unusual anthocyanin reaction with acetone leading to pyranoan-
thocyanin formation. Tetrahedron Letters, 42 (7), 1371–1373.
References
35
© The Author(s) 2016
M. Riaz et al., Anthocyanins and Human Health, SpringerBriefs in Food,
Health, and Nutrition, DOI 10.1007/978-3-319-26456-1_3
Chapter 3
Occurrence of Anthocyanins in Plants
3.1 Introduction
Currently, there are plenty of scientifi c studies dealing with biological and pharma-
cological properties of anthocyanins as well as with their structure, composition,
and abundance in the plant kingdom. Anthocyanins, are considered the most impor-
tant group of fl avonoids in plants having more than 600 compounds identifi ed in
nature. These water-soluble compounds are widely distributed in plant tissues and
provide color to leaves, stems, roots, fl owers and fruits ranging from red, purple to
blue according to the environmental pH and their chemical structure. Due to their
chemical structure anthocyanins are soluble in water solutions and frequently found
in the vacuoles of epidermal cells, and in some species, they are bound to membrane
of the main cell-vacuole, called anthocyanoplasts. The relative abundance, compo-
sition, and chemical structure of anthocyanins may vary according to the species or
from fruit to fruit of the same species depending on external and internal dynamics.
Genetic factors and agronomic practices, intensity and type of light, temperature,
processing season, cultivation and horticultural practices, stage of maturity at har-
vest and post-harvest storage conditions. Thus identifying absolute anthocyanins
identity and content of specifi c plant-derived diet is very diffi cult and therefore
range of values is used usually. Figure 3.1 shows anthocyanins-based colors of
various storage conditions infl uence the level of anthocyanins. Although, anthocya-
nins are found in almost all members of the plant kingdom, plants belonging to 27
families are rich in anthocyanins [ 1 ].
Rich edible sources of anthocyanins are colored fruits such as berries, cherries,
peaches, grapes, pomegranates, and plums as well as many dark-colored vegetables
such as black currant, red onion, red radish, black bean, eggplant, purple corn, red
cabbage, and purple sweet potato [ 2 ]. All these fruits and vegetables are regularly
consumed either in diets or in juices, soft drinks, alcoholic beverages, and similar
other products [ 3 ]. Figure 3.2 summarizes the main known biological functions of
anthocyanins.

36
Almost all highly pigmented fruits and their products as well as byproducts are
potential sources of anthocyanins colorants, such as cranberry press cake, red- grapes
extracts and their by-products [ 4 ], blueberries [ 5 ], elderberries, Hibiscus calyces ,
black choke berries, purple corn, and black current berries etc. However, the antho-
cyanins composition of all of these comprises chiefl y mono and di- glucosides which
stabilize against pH changes and hydration in a limited manner [ 6 ].
Fig. 3.1 Examples of anthocyanins colored fruits, vegetable, leaves and fl owers
3 Occurrence of Anthocyanins in Plants
37
As discussed in Chap. 1 , anthocyanins impart shiny blue, orange, violet, pink and
red colors (Fig. 3.1 ) to the fl owers, vegetables, beverages and fruits of some plants
thus these less stable and more water soluble metabolites than carotenoids, can be
extracted from tulips, roses and orchids [ 7 ]. In addition, Heliophila coronopifolia
from cape areas has blue fl owers [ 8 ] and ‘Dalicha’ ( Tali camellia ) from Yunnan has
red fl owers because of anthocyanins [ 9 ] similarly Rosa hybrida , ( Rosaceae ), has
colored appearance because of anthocyanins and carotenoids [ 10 , 11 ]. Saito et al.
(2011) reported 44 fl oral anthocyanidin glycosides of pelargonidin, cyanidin and
peonidin from the Japanese morning glory ( Ipomoea nil or Pharbitis nil ) [ 8 ].
3.2 Concentration of Anthocyanins in Fruits,
Vegetable and Nuts
As mentioned above, contents and composition of anthocyanins in plant tissue are
affected by genetics (variety and cultivar), geographic origin, growing seasons,
agronomic practices utilized during farming, collection time, climate (temperature,
light, altitude) and geological conditions of area from where that food commodity
or plant part (vegetable, fruit or fl ower) is collected, processing method and the
Functions of
anthocyanins
Protection
against UVB
radiations
Osmotic
adjusters
during
drought or ↓
temperature
Attracting
pollinators
and seed
dispensers
Color in
plants
Light
attenuators
Transport
vehicles for
monosacc-
haride
Active
defense
against fungi
and
herbivores
Fig. 3.2 Functions of anthocyanins in plants
3.2 Concentration of Anthocyanins in Fruits, Vegetable and Nuts
38
method used for determination. Knowledge on individual and population level
anthocyanins intake relies on precise information about their level in foods.
Therefore source and content of anthocyanins in food is of vital importance for
researcher, nutrition-counselors, food processors, manufacturers and consumers.
Several tables and databases of anthocyanins content of foods have been compiled
to facilitate anthocyanins classifi cation and quantifi cation of intake (Tables 3.1 and
3.2 ). Due to the important role in food industry and human nutrition, the common
food of the United States, fruit, vegetable and nuts are assayed for total anthocya-
nins content per 100 g of fresh weight or form consumed (Table 3.1 ).
3.2.1 Variation in Anthocyanins Content
Anthocyanins are present at cellular level in vacuoles, at organ level in fl owers epi-
dermal cells and occasionally mesophylls, however in certain leaves they are
restricted to mesophylls only e.g. leaves of Secale cereal [ 40 ]. Different food matri-
ces require different methods for complete release of anthocyanins without degrada-
tion and therefore accurate determination of anthocyanins contents requires
optimum and validated procedures both for isolation and quantifi cation. Even for
same food matrix, various factor like varietal differences, time of harvest, farming
practices and raw versus processed food infl uence the anthocyanin contents.
Sometimes many of above mentioned variables affect anthocyanin contents and
Table 3.1 Anthocyanins content of commonly consumed fruits and vegetables in United States [ 12 ]
Fruit
Total
anthocyanins (mg) Fruit
Total
anthocyanins (mg)
Apple (Fuji, Gala, Red
delicious)
1.8, 3.2 and 17 Marion blackberry 433
Blackberry 353 Blueberry cultivated 529
Gooseberry (average of
various cultivars)
6.43 Elderberry 1993
Chokeberry 2147 Cranberry 133
Peach 4.7 Concord grape 192
Nectarine 9.2 Red grape 42.7
Currant black, 533 Currant red 14.3
Cherry (sweet) 177 Blueberry wild 705
Strawberry (average) 52.1 Raspberry black,
red
845, 116
Black plum 82 Plum 12.5
Blackbeans 23.1 Red cabbage 113
Eggplant 35.1 Red leaf lettuce 1.5
Red onion 38.8 Small red bean 6.2
Red radish 116 Pistachio 2.1
mg per 100 g fresh weight
3 Occurrence of Anthocyanins in Plants
39
while it may be possible to make generalization about one factor, in reality numer-
ous factors exert their infl uence during synthesis and accumulation of anthocyanins.
Further their post-harvest stability is affected by processing, cooking, heat-
treatment, exposure to light and oxygen and storage which not only affects content
but also form of anthocyanins. The concentration of anthocyanins is variable, it was
reported to be 0.1–1 % per dry weight for most of the fruits and vegetable, it may be
present as single main type e.g. Dianthus, apple, cherry, two main type e.g. cranberry
and many/mixture of anthocyanins e.g. Rosa , Tulipa , grapes [ 40 ]. Table 3.3 shows
various rich sources of anthocyanins.
Andersen and Jordheim (2006) reported that there are more than 500 different
anthocyanins [ 47 ] and 23 anthocyanidins [ 47 , 48 ]; of which only six are the most
common in vascular plants, pelargonidin, peonidin, cyanidin, malvidin, petunidin
and dephinidin [ 49 ]. The glycoside derivatives of the cyanidin, dephinidin and
pelargonidin, i.e. three non-methylated anthocyanidins, are mostly found in nature,
with 50 %,69 % and 80 % in fl owers, fruits and leaves respectively [ 50 ].
The distribution of the six common anthocyanidins in fruits and vegetables in the
abundance order as: cyanidin (50 %), pelargonidin (12 %), peonidin (12 %), delph-
inidin (12 %), petunidin (7 %), and malvinidin (7 %), while publishing another
report he included more anthocyanins sources and the abundance order was likely
to be cyanidin (30 %), delphinidin (22 %), pelargonidin (18 %), peonidin (7.5 %),
malvidin (7.5 %), and peunidin (5 %). In both reports, the three non-methylated
anthocyanidins (cyanidin, delphinidin, and pelargonidin) were exposed to be more
prevalent than the three methylated anthocyanins (peonidin, malvidin, and petuni-
din). So it was confi rmed that cyanidin-3-glucoside is the most common anthocya-
Table 3.2 Plants as sources of anthocyanins
Anthocyanins sources Reference Anthocyanins sources Reference
Vegetables [
13 ] Elderberries [ 14 ]
Potato [
15 ] Hibiscus calyces [ 16 ]
Tomato [
17 ] Black currents [ 18 ]
Spinach [
19 ] Purple corn [ 20 ]
Paprika [
21 ] Berries [ 22 ]
Purple corn [
23 ] Strawberry [ 24 ]
Red radish [
25 ] Mulberry [ 26 ]
Red and green cabbages [
27 , 28 ] Tamarillo fruit ( Solanum
betaceum Cav.)
[ 29 ]
Fruits Garcinia indica Choisy [
30 ]
Cherries [
31 ] Litchi [ 32 ]
Extracts of red grapes and its
by-products
[
33 ] Apple skin [ 34 ]
Cranberry press cake [
35 , 36 ] Pomegranate
( Punica granatum L.)
[ 37 ]
Blueberries [
5 ] Muscadine grapes
( Vitis rotundifolia )
[ 38 ]
Black chokeberries [
39 ]
3.2 Concentration of Anthocyanins in Fruits, Vegetable and Nuts
40
nins in nature and more than 90 % of anthocyanins contain glucose as a glycosylating
sugar [ 7 , 40 ].
3.3 Typical New Anthocyanins Found in the Past Years
Four anthocyanins with furanose sugar (apiose) were obtained from leaves of
Synadenium grantii ( Euphorbiaceae ) [ 51 ], which is very unusual in anthocyanin
family. In 2003 and 2004, Honda and Tatsuzawa et al. (2004) discovered two novel
C-glycosyl anthocyanins, which were isolated from the fl owers of Tricyrtis formo-
sana cultivar Fujimusume [ 52 ]. Pyranoanthocyanins are a newer member with a
pyran ring in the anthocyanins family, which were reported in 1996 as the pigments
detected in red wines fi ltrates [ 53 ]. Furthermore, there are some unique anthocyanin–
fl avanol condensed pigments (polymeric anthocyanins) reported in existing studies
[ 54 , 55 ]. Table 3.4 shows major and minor anthocyanins in various food while
Table 3.5 shows the occurrence of anthocyanins in common Mediterranean diet.
Table 3.3 Anthocyanins content in vegetables and fruits
Sources Cultivar
Total anthocyanin
content (mg/100 g) Reference
Blackberries, raw Rubus fruticosus 100.61 [
41 ]
Blueberries, raw Vaccinium myrtillus 163.30 [
41 ]
Cranberry, raw Vaccinium oxycoccos 103.70 [
41 ]
Gooseberry, raw Ribesuva - crispa 9.51 [
41 ]
Raspberry, raw Rubus idaeus 48.63 [
41 ]
Strawberry, raw Fragaria xananassa 27.01 [
41 ]
Grape, raw Vitis vinifera 48.04 [
41 ]
Grape, dried Sunbelt 107.60 [
42 ]
Grape, raw, skin Black Olympia 97.50 [
43 ]
Pomegranate, raw Mollar de Elche 56.09 [
43 ]
Purple tomato, dried V118 72.31 [
44 ]
Purple carrot, dried Rain 44.27 [
45 ]
Purple carrot, dried Haze 57.22 [
45 ]
Red cabbage, dried Gario 198.61 [
45 ]
Purple caulifl ower, dried Graffi tti 201.11 [
45 ]
Purple potato, dried Majesty 97.71 [
45 ]
Purple potato, dried Mackintosh 48.74 [
45 ]
Red potato, dried Y38 85.23 [
45 ]
Red potato, dried Thumb 43.09 [
45 ]
Red onion, dried Pier - c 7.77 [
45 ]
Red onion, dried Pearl 18.95 [
45 ]
Red onion, raw Morada de Amposta 23.30 [
46 ]
Eggplant, dried Black Beauty 29.55 [
45 ]
3 Occurrence of Anthocyanins in Plants
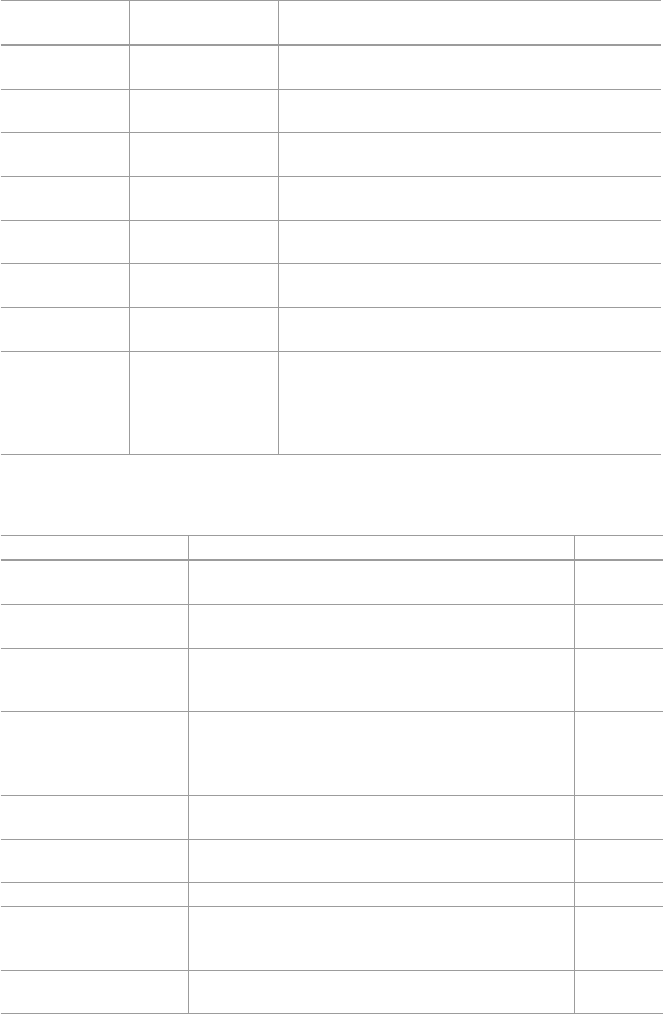
41
Table 3.4 Occurrence pattern of anthocyanins in common vegetables and fruits [
56 ]
Name
Major
anthocyanins Minor anthocyanins
Strawberry Pelargonidin-
3- glucoside
Cyanidin-3-glucoside, Pelargonidin-3-rutinoside
Blackberry Cyanidin-
3- glucoside
Cyanidin-3-rutinoside, malvidin-3-glucoside
Raspberry Cyanidin-
3- glucoside
Pelargonidin-3-glucosides, Pelargonindin-3-rutinoside
Sweet cherries Cyanidin-
3- rutinoside
Cyanidin-3-glucoside, Peonidin-3-rutinoside
Blackcurrant Cyanindin-
3- rutinoside
Cyanindin-3-glucoside, Delphinidin-3-glucoside
Bilberry Delphinidin-
3- galactoside
Peonindin-3-glucoside, Peonindin-3-galactoside
Red onions Cyanidin
3-glucoside
Delphinidin-3-glucoside, Petunidinglucoside
Blood orange Cyanidin
3-glucoside
Delphinidin 3-glucoside, cyanidin 3,5-diglucoside,
cyanidin 3-sophoroside, delphinidin
3-(6″-malonylglucoside), peonidin
3-(6″-malonylglucoside), cyanidin
3-(6″-dioxalylglucoside)
Table 3.5 The occurrence of anthocyanins in common Mediterranean diet
Plant Anthocyanins References
Blackberries ( Rubus
ulmifolius Schott )
Cyanidin-3-glucoside (major) and cyanidin-3-
rutinoside (minor)
[
57 ]
Eggplant ( Solanumm
elongena L .)
Nasunin [
58 ]
Fig ( Ficus carica L .) Cyanidin-3- O -rhamnoglucoside (cyanidin-3- O -
rutinoside) (major), cyanidin 3-glucoside, cyanidin
3,5-diglucoside and pelargonidin 3-rutinoside
[
59 , 60 ]
Grape ( Vitis vinifera L .)
and Wine
3- O -monoglucoside of pelargodin-3- O -glucoside,
cyanidin-3- O -glucoside, delphinidin-3- O -glucoside,
peonidin-3- O -glucoside, petunidin-3- O -glucosideand
malvidin-3- O -glucoside
[
61 ]
Lentil ( Lens culinaris
Medik .)
Delphinidin-3- O -(2- O - β-
D -glucopyranosyl-α- L -
arabino- pyranoside
[ 62 ]
Myrtle ( Myrtus
communis L .)
Anthocyanin arabinosides [
63 ]
Olive ( Olea europea L .) Cyanidin 3- O -glucoside and cyanidin 3- O -rutinoside [
64 ]
Sweet Cherry ( Prunu
savium L .)
Cyanidin 3-rutinoside, cyanidin 3-glucoside, peonidin
3-rutinoside, cyanidin 3-sophoroside, pelargonidin
3-glucoside, pelargonidin 3-rutinoside, 3-glucoside
[
65 , 66 ]
Plum ( Prunus
domestica L .)
Cyanidin-3-glucoside, cyanidin-3-rutinoside,
peonidin- 3- glucoside, and peonidin-3-rutinoside
[
67 , 68 ]
(continued)
3.3 Typical New Anthocyanins Found in the Past Years
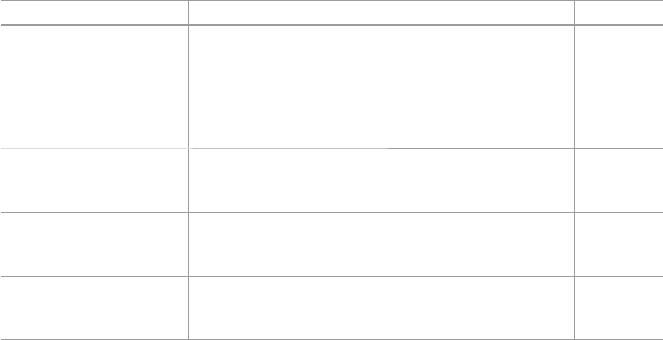
42
3.4 Conclusions
Various fruits, vegetables, nuts, herbs, spices and horticultural crops are good source
of anthocyanins. Diverse factors like packaging, freezing, lipid composition and
other bioactive constituents present in fruits and vegetables affect anthocyanin
contents. Release of organic acids that can lead to isomerization during slicing and
juicing of fruits, surface area, porosity and enzymatic oxidation during slicing,
peeling, pulping or juicing all affect anthocyanins contents in fruits. Therefore
marketing minimally processed fruits and vegetables is continuously increasing due
to consumer demand for high quality and nutritive products. The development of
novel and more ample sources of anthocyanins in horticultural crops, vegetables
and fruits and improvement of their shelf-life and consumer acceptance can improve
human health signifi cantly. The ongoing research on horticulture and crops is
expected to bring more verities on grocery shelves and in hands of consumers
having more anthocyanins contents than traditional varieties.
References
1. Ghosh, D., & Konishi, T. (2007). Anthocyanins and anthocyanin-rich extracts: Role in diabetes
and eye function. Asia Pacifi c Journal of Clinical Nutrition, 16 (2), 200.
2. Wu, X., Pittman, H. E., & Prior, R. L. (2006). Fate of anthocyanins and antioxidant capacity in
contents of the gastrointestinal tract of weanling pigs following black raspberry consumption.
Journal of Agricultural and Food Chemistry, 54 (2), 583–589.
3. Valls, J., Millán, S., Martí, M. P., Borràs, E., & Arola, L. (2009). Advanced separation methods
of food anthocyanins, isofl avones and fl avanols. Journal of Chromatography A, 1216 (43),
7143–7172.
Plant Anthocyanins References
Onion ( Allium cepa L .) 3-(3-Glucosyl-6-malonyl-glucoside), 3-(6-malonyl-
glucoside), 3-(3-glucosylglucoside) and cyanidin-3-
glucoside, respectively 3-(3,6-dimalonylglucoside),
3-(3-malonylglucoside), and 3,5-diglucoside of
cyanidin, 3-glucoside, 3,5-diglucoside and
peonidin-3-malonyl-glucoside
[
69 , 70 ]
Red Radish ( Raphanus
sativus L .)
Cyanidin and pelargonidin derivatives pelargonidin-3-
sophoroside-5-glucoside derivative compounds were
found
[
71 ]
Red chicory ( Cichorium
intybus L. var. Silvestre
Bisch .)
Cyanidin-3- O -(6-malonyl-β-glucopyranoside)
delphinidin derivative has been recently identifi ed
[
72 , 73 ]
Pomegranate ( Punica
granatum L .)
3-Glucosides and 3,5-diglucoside such as
delphinidin-3- glucoside, cyanidin-3,5-diglucoside,
delphinidin-3- glucoside and pelargonidin-3-glucoside
[
74 , 75 ]
Table 3.5 (continued)
3 Occurrence of Anthocyanins in Plants
43
4. Camire, A., & Clydesdale, F. (1979). High pressure liquid chromatography of cranberry antho-
cyanin. Journal of Food Science, 44 (3), 926–927.
5. Francis, F. (1985). Blueberries as a colorant ingredient in food products. Journal of Food
Science, 50 (3), 754–756.
6. Giusti, M. M., & Wrolstad, R. E. (2003). Acylated anthocyanins from edible sources and their
applications in food systems. Biochemical Engineering Journal, 14 (3), 217–225.
7. Castañeda-Ovando, A., Pacheco-Hernández, M., Páez-Hernández, M. E., Rodríguez, J. A., &
Galán-Vidal, C. A. (2009). Chemical studies of anthocyanins: A review. Food Chemistry,
113 (4), 859–871.
8. Saito, N., Tatsuzawa, F., Toki, K., Shinoda, K., Shigihara, A., & Honda, T. (2011). The blue
anthocyanin pigments from the blue fl owers of Heliophila coronopifolia L. (Brassicaceae).
Phytochemistry, 72 (17), 2219–2229.
9. Li, J. B., Hashimoto, F., Shimizu, K., & Sakata, Y. (2008). Anthocyanins from red fl owers of
Camellia cultivar ‘Dalicha’. Phytochemistry, 69 (18), 3166–3171.
10. Lee, J. H., & Choung, M.-G. (2011). Identifi cation and characterisation of anthocyanins in the
antioxidant activity-containing fraction of Liriope platyphylla fruits. Food Chemistry, 127 (4),
1686–1693.
11. Lee, J. H., Lee, H.-J., & Choung, M.-G. (2011). Anthocyanin compositions and biological
activities from the red petals of Korean edible rose ( Rosa hybrida cv. Noblered). Food
Chemistry, 129 (2), 272–278.
12. Wu, X., Beecher, G. R., Holden, J. M., Haytowitz, D. B., Gebhardt, S. E., & Prior, R. L. (2006).
Concentrations of anthocyanins in common foods in the United States and estimation of nor-
mal consumption. Journal of Agricultural and Food Chemistry, 54 (11), 4069–4075.
13. Lutz, M., Hernández, J., & Henriquez, C. (2015). Phenolic content and antioxidant capacity in
fresh and dry fruits and vegetables grown in Chile. CyTA Journal of Food, 13 (4), 541–547.
14. Brønnum Hansen, K., & Flink, J. (1986). Anthocyanin colorants from Elderberry ( Sambucus
nigra L.) IV. Further studies on production of liquid extracts, concentrates and freeze dried
powders. International Journal of Food Science and Technology, 21 (2), 605–614.
15. Friedman, M. (1997). Chemistry, biochemistry, and dietary role of potato polyphenols.
A review. Journal of Agricultural and Food Chemistry, 45 (5), 1523–1540.
16. Pouget, M. P., Lejeune, B., Vennat, B., & Pourrat, A. (1990). Extraction, analysis and study of
the stability of Hibiscus anthocyanins. Lebensmittel–Wissenschaft and Technology, 23 (2),
103–105.
17. Abdulnabi, A., Abushita, E. A., Hebshi, H. G. D., & Biacs, P. A. (1997). Determination of
antioxidant vitamins in tomatoes. Food Chemistry, 60 (2), 207–212.
18. Rosa, J. (1973). Stability of anthocyanin pigment concentrates obtained from black current
press cake. II. Studies on the rate of pigment destruction during storage. Prace Instytut6w i
Laboratoridw Badawczych Przemyslu Spolywczego, 23 (3), 447–462.
19. Gil, M. I., Ferreres, F., & Tomas-Barberan, F. A. (1999). Effect of postharvest storage and
processing on the antioxidant constituents (fl avonoids and vitamin C) of fresh-cut spinach.
Journal of Agricultural and Food Chemistry, 47 (6), 2213–2217.
20. Nakatani, N., Fukuda, H., & Fuwa, H. (1979). Major anthocyanin of Bolivian purple corn ( Zea
mays L.). Agri Biol Chem, 43 (2), 389–391.
21. Márkus, F., Daood, H. G., Kapitány, J., & Biacs, P. A. (1999). Change in the carotenoid and
antioxidant content of spice red pepper (paprika) as a function of ripening and some techno-
logical factors. Journal of Agricultural and Food Chemistry, 47 (1), 100–107.
22. Li, R., Wang, P., Qing-qi, G., & Zhen-yu, W. (2011). Anthocyanin composition and content of
the Vaccinium uliginosum berry. Food Chemistry, 125 (1), 116–120.
23. Yang, Z., Yonbin, H., Zhenxin, G., Gongjian, F., & Zhigang, C. (2008). Thermal degradation
kinetics of aqueous anthocyanins and visual color of purple corn ( Zea mays L.) cob. Innovative
Food Science and Emerging, 9 (3), 341–347.
24. Bordonaba, J. G., Crespo, P., & Terry, L. A. (2011). A new acetonitrile-free mobile phase for
HPLC-DAD determination of individual anthocyanins in blackcurrant and strawberry fruits:
A comparison and validation study. Food Chemistry, 129 (3), 1265–1273.
References
44
25. Patil, G., Madhusudhan, M. C., Ravindra, B. B., & Raghavarao, K. S. M. S. (2009). Extraction,
dealcoholization and concentration of anthocyanin from red radish. Chemical Engineering and
Processing, 48 (1), 364–369.
26. Du, Q., Zheng, J., & Xu, Y. (2008). Composition of anthocyanins in mulberry and their anti-
oxidant activity. Journal of Food Composition and Analysis, 21 (5), 390–395.
27. Arapitsas, P., & Turner, C. (2008). Pressurized solvent extraction and monolithic column-
HPLC/DAD analysis of anthocyanins in red cabbage. Talanta, 74 (5), 1218–1223.
28. McDougall, G. J., Fyffe, S., Dobson, P., & Stewart, D. (2007). Anthocyanins from red cab-
bage-stability to simulated gastrointestinal digestion. Phytochemistry, 68 (9), 1285–1294.
29. Nelson, H., Hurtado, A. L., Morales, M., González-Miret, L., Escudero-Gilete, M. L., &
Francisco, J. H. (2009). Colour, pH stability and antioxidant activity of anthocyanin rutino-
sides isolated from tamarillo fruit ( Solanum betaceum Cav.). Food Chemistry, 117 (1), 88–93.
30. Nayak, C. A., Rastogi, N. K., & Raghavarao, K. (2010). Bioactive constituents present in
Garcinia indica Choisy and its potential food applications: A review. International Journal of
Food Properties, 13 (3), 441–453.
31. Ara Kirakosyan, E., Mitchell, S., Kathleen, R., Noon, D. E., Urcuyo, L., Peter, B., et al. (2010).
Interactions of antioxidants isolated from tart cherry ( Prunus cerasus ) fruits. Food Chemistry,
122 (1), 78–83.
32. Ara Kirakosyan, E., Mitchell, S., Kathleen, R., Noon, D. E., Urcuyo, L., Peter, B., et al. (2010).
Interactions of antioxidants isolated from tart cherry ( Prunus cerasus ) fruits. Food Chemistry,
122 (1), 78–83.
33. Markakis, P. (2012). Anthocyanins as food colors . New York: Elsevier.
34. Ubi, B. E., Honda, C., Bessho, H., Kondo, S., Wada, M., Kobayashi, S., et al. (2006).
Expression analysis of anthocyanin biosynthetic genes in apple skin: Effect of UV-B and tem-
perature. Plant Science, 170 (3), 571–578.
35. Sapers, G., Taffer, I., & Ross, L. (1981). Functional properties of a food colorant prepared
from red cabbage. Journal of Food Science, 46 (1), 105–109.
36. Chiriboga, C., & Francis, F. (1970). Anthocyanin recovery system from cranberry pomace.
Proceedings of the American Society for Horticultural Science, 95 (2), 233–236.
37. Turfan, O., Türkyılmaz, M., Yemiş, O., & Özkan, M. (2011). Anthocyanin and colour changes
during processing of pomegranate ( Punica granatum L., cv. Hicaznar) juice from sacs and
whole fruit. Food Chemistry, 129 (4), 1644–1651.
38. Huang, Z., Baowu, W., Paul, W., & Ralphenia, D. P. (2009). Identifi cation of anthocyanins in
muscadine grapes with HPLC-ESI-MS. LWT – Food Science and Technology, 42 (4), 819–824.
39. Kraemer-Schafhalter, A., Fuchs, H., & Pfannhauser, W. (1996). Anthocyanins of aronia mela-
nocarpa: Analysis, stability, changes during treatment and storage. In: Proceedings of the sym-
posium on Polyphenols and anthocyanins as food colorants and antioxidants, Vienna, 15 Nov
1996.
40. Delgado-Vargas, F., Jiménez, A., & Paredes-López, O. (2000). Natural pigments: Carotenoids,
anthocyanins, and betalains—Characteristics, biosynthesis, processing, and stability. Critical
Reviews in Food Science and Nutrition, 40 (3), 173–289.
41. Kruger, M. J., Neil, D., Myburgh, K. H., & Sandrine, L. (2014). Proanthocyanidins, anthocya-
nins and cardiovascular diseases. Food Research International, 59 , 41–52.
42. Khanal, R. C., Howard, L. R., & Prior, R. L. (2010). Effect of heating on the stability of grape
and blueberry pomace procyanidins and total anthocyanins. Food Research International,
43 (5), 1464–1469.
43. Zhenchang, L., Benhong, W., Peige, F., Chunxiang, Y., Wei, D., Xianbo, Z., et al. (2008).
Anthocyanin composition and content in grape berry skin in Vitis germplasm. Food Chemistry,
111 (4), 837–844.
44. Li, H., Deng, Z., Liu, R., Young, J. C., Zhu, H., Loewen, S., et al. (2011). Characterization of
phytochemicals and antioxidant activities of a purple tomato ( Solanum lycopersicum L.).
Journal of Agricultural and Food Chemistry, 59 (21), 11803–11811.
45. Li, H., Zeyuan, D., Honghui, Z., Chanli, H., Ronghua, L. J., Christopher, Y., et al. (2012).
Highly pigmented vegetables: Anthocyanin compositions and their role in antioxidant activi-
ties. Food Research International, 46 (1), 250–259.
3 Occurrence of Anthocyanins in Plants
45
46. Ferreres, F., Gil, M. I., & Tomás-Barberán, F. A. (1996). Anthocyanins and fl avonoids from
shredded red onion and changes during storage in perforated fi lms. Food Research International,
29 (3), 389–395.
47. Anderson, O., & Jordheim, M. (2006). The anthocyanins. In O. M. Anderson & K. R.
Markhand (Eds.), Flavonoids chemistry, biochemistry and application . Boca Raton: CRC.
48. Rein, M. (2005). Copigmentation reactions and color stability of berry anthocyanins . Helsinki:
University of Helsinki.
49. Clifford, M. N. (2000). Anthocyanins-nature, occurrence and dietary burden. Journal of the
Science of Food and Agriculture, 80 (7), 1063–1072.
50. Dey, P. M., & Harborne, J. B. (1997). Plant biochemistry . New York: Academic.
51. Andersen, O. M., Jordheim, M., Byamukama, R., Mbabazi, A., Ogweng, G., Skaar, I., et al.
(2010). Anthocyanins with unusual furanose sugar (apiose) from leaves of Synadenium grantii
(Euphorbiaceae). Phytochemistry, 71 (13), 1558–1563.
52. Tatsuzawa, F., Norio, S., Hiroko, S., Masato, Y., Tomohisa, Y., Koichi, S., et al. (2004).
Acylated anthocyanins in the fl owers of Vanda (Orchidaceae). Biochemical Systematics and
Ecology, 32 (7), 651–664.
53. Cameira dos Santos, P. J., Jean-Marc, B., Véronique, C., & Michel, M. (1996). Detection and
partial characterisation of new anthocyanin derived pigments in wine. Journal of the Science
of Food and Agriculture, 70 (2), 204–208.
54. Glenda, A., Macz-Pop, J. C., Rivas-Gonzalo, J., Pérez-Alonso, J., & Ana, M. G. P. (2006).
Natural occurrence of free anthocyanin aglycones in beans ( Phaseolus vulgaris L.). Food
Chemistry, 94 (3), 448–456.
55. João, P., Sandra, L., Ana, M. G. P., Nuno, M. C., Santos, B., Artur, M. S. S., et al. (2005).
Isolation and structural characterization of new anthocyanin-alkyl-catechin pigments. Food
Chemistry, 90 (1), 81–87.
56. Patras, A., Nigel, P. B., Colm, O. D., & Tiwari, B. K. (2010). Effect of thermal processing on
anthocyanin stability in foods; mechanisms and kinetics of degradation. Trends in Food
Science and Technology, 21 (1), 3–11.
57. Fan‐Chiang, H. J., & Wrolstad, R. E. (2005). Anthocyanin pigment composition of blackber-
ries. Journal of Food Science, 70 (3), C198–C202.
58. Kuroda, C., & WADA, M. (1933). The colouring matter of eggplant (Nasu 1). Proceedings of
the Imperial Academy, 9 (2), 51–52.
59. Solomon, A., Golubowicz, S., Yablowicz, Z., Grossman, S., Bergman, M., Gottlieb, H. E., et
al. (2006). Antioxidant activities and anthocyanin content of fresh fruits of common fi g ( Ficus
carica L.). Journal of Agricultural and Food Chemistry, 54 (20), 7717–7723.
60. Dueñas, M., José, J. P., Santos-Buelga, C., & Escribano-Bailón, T. (2008). Anthocyanin com-
position in fi g ( Ficus carica L.). Journal of Food Composition and Analysis, 21 (2), 107–115.
61. Warner, L. M. (Ed.). (2015). Handbook of anthocyanins: Food sources, chemical applications
and health benefi ts . New York: Nova Science.
62. Takeoka, G. R., Dao, L. T., Tamura, H., & Harden, L. A. (2005). Delphinidin 3-O-(2-O-β-D-
glucopyranosyl-α-L-arabinopyranoside): A novel anthocyanin identifi ed in beluga black len-
tils. Journal of Agricultural and Food Chemistry, 53 (12), 4932–4937.
63. Montoro, P., Tuberoso, C. I., Perrone, A., Piacente, S., Cabras, P., & Pizza, C. (2006).
Characterisation by liquid chromatography-electrospray tandem mass spectrometry of antho-
cyanins in extracts of Myrtus communis L. berries used for the preparation of myrtle liqueur.
Journal of Chromatography A, 1112 (1), 232–240.
64. Aparicio-Ruiz, R. N., Gandul-Rojas, B., & Roca, M. (2009). Pigment profi le in non-Spanish
olive varieties ( Olea europaea L. Var. Coratina, Frantoio, and Koroneiki). Journal of
Agricultural and Food Chemistry, 57 (22), 10831–10836.
65. Grigoras, C. G., Emilie, D., Sandrine, Z., & Claire, E. (2012). Sweet cherries anthocyanins: An
environmental friendly extraction and purifi cation method. Separation and Purifi cation
Technology, 100 , 51–58.
66. Usenik, V., Fabčič, J., & Štampar, F. (2008). Sugars, organic acids, phenolic composition and
antioxidant activity of sweet cherry ( Prunus avium L.). Food Chemistry, 107 (1), 185–192.
References
46
67. Kim, D. O., Chun, O. K., Kim, Y. J., Moon, H. Y., & Lee, C. Y. (2003). Quantifi cation of poly-
phenolics and their antioxidant capacity in fresh plums. Journal of Agricultural and Food
Chemistry, 51 (22), 6509–6515.
68. Chun, O. K., Kim, D.-O., & Lee, C. Y. (2003). Superoxide radical scavenging activity of the
major polyphenols in fresh plums. Journal of Agricultural and Food Chemistry, 51 (27),
8067–8072.
69. Fossen, T., Andersen, O. M., Dag, O., Ovstedal, A. T., & Pedersen, A. R. (1996). Characteristic
anthocyanin pattern from onions and other Allium spp. Journal of Food Science, 61 (4),
703–706.
70. Donner, H., Gao, L., & Mazza, G. (1997). Separation and characterization of simple and malo-
nylated anthocyanins in red onions, Allium cepa L. Food Research International, 30 (8),
637–643.
71. Jing, P., Shu-Juan, Z., Si-Yu, R., Zhuo-Hong, X., Ying, D., & Liangli, Y. (2012). Anthocyanin
and glucosinolate occurrences in the roots of Chinese red radish ( Raphanus sativus L.), and
their stability to heat and pH. Food Chemistry, 133 (4), 1569–1576.
72. Carazzone, C., Mascherpa, D., Gazzani, G., & Papetti, A. (2013). Identifi cation of phenolic
constituents in red chicory salads ( Cichorium intybus ) by high-performance liquid chromatog-
raphy with diode array detection and electrospray ionisation tandem mass spectrometry. Food
Chemistry, 138 (2), 1062–1071.
73. Mulabagal, V., Haibo, W., Mathieu, N., & Muraleedharan, G. N. (2009). Characterization and
quantifi cation of health benefi cial anthocyanins in leaf chicory ( Cichorium intybus ) varieties.
European Food Research and Technology, 230 (1), 47–53.
74. Mena, P., Calani, L., Dall’Asta, C., Galaverna, G., García-Viguera, C., Bruni, R., et al. (2012).
Rapid and comprehensive evaluation of (poly) phenolic compounds in pomegranate ( Punica
granatum L.) juice by UHPLC-MSn. Molecules, 17 (12), 14821–14840.
75. Zhao, X., Zhaohe, Y., Fang, Y., Yanlei, Y., & Lijuan, F. (2013). Characterization and evaluation
of major anthocyanins in pomegranate ( Punica granatum L.) peel of different cultivars and
their development phases. European Food Research and Technology, 236 (1), 109–117.
3 Occurrence of Anthocyanins in Plants
47
© The Author(s) 2016
M. Riaz et al., Anthocyanins and Human Health, SpringerBriefs in Food,
Health, and Nutrition, DOI 10.1007/978-3-319-26456-1_4
Chapter 4
Anthocyanins as Natural Colors
4.1 Introduction
Colors play a very important but often under-estimated role in our lives. Since
ancient times natural colors derived from plants, lichens and insects have been used
as sources for natural colors, suitable for multiple applications including textiles,
paints, works of art, food, clothing and cosmetics. Colors fascinate human and ani-
mals as they are one of the fi rst characteristics observed by senses and helps in
determining the acceptability of food and play a crucial role in the plant pollination
process by animals.
Since, a decent outlook is crucial to fetch high market price of food item, natural
or synthetic color is added to the food. Natural pigments from plant or animal origin
are believed to be more safe and cost-effective than synthetic ones. Synthetic dyes
have been suspected to affect adversely central nervous system both behaviorally
and neurologically. Colors from biological origin are called ‘biocolors’. The suc-
cess of any color, natural or synthetic, depends upon its consumer-acceptance,
approval by regulatory bodies and the size of the investment required in bringing the
color to store-shelves. The use of natural colors is an infl uential tool for selling a
product in market. The food as such or in packed form is judged by its color or even
by color of package. It has led to claims appearing on colored food products and
commodities like, “no synthetic added” or “all natural”.
There are many pigmented classes of phytochemicals, such as carotenoids, fl avo-
noids and anthocyanins. The later class has been focus of attention as these are safe
compared to artifi cial colors like Sudan red III and other synthetic colors.
Anthocyanins like other bio-colors have a high market value with prices variations
from US $1250 to $2000 per kg. No permission is required for work on these pig-
ments as these occur naturally.
Chlorophyll, carotenoids, and anthocyanins are the most abundant natural occur-
ring pigments in plant tissues. We recognize anthocyanins indirectly because of the
splendid red to red-brown to purple autumn colors of the leaves of trees. In combi-
48
nation with the yellow red carotenoids, they give leaves, fl owers and fruits a wide
spectrum of colors. The anthocyanins are now exclusively used as food colors but in
older times they were used in paints for precious manuscripts. They are now pre-
pared from the peels of grapes (enocyanin) as a side product of the grape harvest
and winemaking. Red cabbage is also a good source of anthocyanins. The rainbow
of colors of anthocyanins is due to conjugated bonds present in their structures.
Most of the fl ower colors are fl avonoids in nature and red or purple is associated
with anthocyanins alone or as co-pigment responsible for other related colors. The
pigmentation of fl ower attract insects and contribute in its propagation, another
important role of anthocyanins for plants is protection from external plant enemies
e.g. cyanidin and peonidin glucosides inhibited the growth of Xanthomonas oryzae
[ 1 ]. Their importance may not be overlooked because they have been sub-grouped
as color of current EU approved additives with E163, E-Number by EU food stan-
dard agency [ 2 ]. Safety and water solubility of anthocyanins allow their integration
into aqueous foodstuff without unwanted negative effects [ 3 ].
Like synthetic color, pure single natural color is hard to obtain thus extract from
the concerned natural source is used as coloring agent for example the most com-
monly anthocyanins enriched extract used are grape peel and black currant. This
natural color varies with the stage and season of the plant, leaves when young have
different color than aged leaves and same is for fruit and fl owers. The reason for
color variation is thought to be the intra and intermolecular bioconversion of antho-
cyanins or derivatives with age and environment to its variant analog moieties that
have specifi c colors. For example the interconversion of aromatic polyacylated
anthocyanins is responsible for bluing effect and stability of fl ower color. Taking the
advantage of the advancement in plant biotechnology the cultivar of desired color
may be produced by incorporating the gene of interest. Investigation on this side is
required to produce natural color in greater in quantity and quality.
As mentioned above, the color of anthocyanins is due to the conjugated double
bonds carrying a positive charge on the heterocyclic oxygen ring under acidic con-
ditions. Various factors like pH, co-pigmentation and interactions with metal ions
lead to variation of colors in anthocyanins by affecting the conjugated double bond
system present in basic skeleton of anthocyanins. At pH near to 1 the free fl avylium
cation is red while at higher pH, the color changes to purple and blue as it goes in
transition to the quinoidal base and subsequently to ionized quinoidal base. At
higher pH, the colorless carbinol pseudo-base is formed which ultimately changes
to chalcone molecule having a pale yellow color. Substitution of hydroxyl or methyl
group also affect the color of anthocyanins (Figs. 4.1 and 4.2 ) for example increas-
ing the number of hydroxyl increase bluishness, while increasing the number of
methyl group intensify redness [ 1 ].
Anthocynanin color depends upon number of OH groups attached especially on
B ring. Addition of OH groups shifts their color from orange to violet. Similarly
glycosylation of anthocyanidin leads to extra red color of anthocyanins while pres-
ence of aromatic or aliphatic acyl moieties does not change color but affects antho-
cyanin solubility as well as stability. Literature indicates that more than 650
anthocyanins exist in nature. In 1993, over 250 different anthocyanins have been
4 Anthocyanins as Natural Colors
49
OH
H
OO
+
O
H
Apigeninidin
(Orange)
O
+
O
H
O
H
O
H
O
H
H
O
O
H
Delphinidin
(Purple, mauve and blue)
OH
H
OO
+
O
H
OH
H
O
Aurantinidin
(Orange)
OH
H
OO
+
O
H
OH
O
H
Cyanidin
(Magenta and Crimson)
OH
H
OO
+
O
H
OH
HO
O
H
6-Hydroxycyanidin
(Red)
OH
H
OO
+
O
H
OH
HO
O
H
Luteolinidin
(Orange)
O
+
O
H
OH
OH
H
O
Pelargonidin
(Orange, salmon)
O
+
O
H
OH
OH
H
O
Triacetidin
(Red)
O
H
OH
Fig. 4.1 Natural OH substituent’s anthocyanidins and change in color [ 1 ]
4.1 Introduction
50
O
+
OH
O
OH
OH
H
O
Peonidin
(Magenta)
Capensinidin
(Bluish red)
O
+
OH
O
OH
O
HO
O
Europenidin
(Bluish red)
O
+
OH
OH
O
H
O
O
O
+
OH
OH
OH
O
O
Hirsutidin
(Bluish red)
O
O
+
OH
O
OH
OH
H
O
O
Malvidin
(Purple)
O
+
O
H
OH
O
H
O
O
H
5-Methylcyanidin
(Orange red)
O
+
O
H
O
OH
OH
H
O
O
H
Petunidin
(Purple)
O
+
O
H
OH
OH
O
H
O
O
H
Pulchellidin
(Bluish red)
O
+
OH
OH
OH
O
O
Rosinidin
(Red)
Fig. 4.2 Natural methyl substituent’s anthocyanidins and change in color [ 1 ]
4 Anthocyanins as Natural Colors
51
isolated from plants [ 4 – 7 ] and 500 anthocyanin structures reported by the year 2000
[ 8 ]. The six common anthocyanidins yield greater than 540 anthocyanins in nature
[ 9 ], due to acylation of sugar residues with organic-acids and structural differences
of glycosidic substitution at the 3 and 5 positions [ 10 ]. Andersen and Jordheim, in
2006 reported the number 500 [ 10 ] and the number increased to 635 in 2008 [ 11 ].
Anthocynains are exclusively used as red color for food stuff although sometime
betalains are also used. The chief reason of this use in food matrixes is requirement
of marinating the acidic pH (less than 3.5) to obtain the required red color. Grape
extracts are more stable towards pH variations so they are widely used as commer-
cial sources of anthocyanins. Further grapes are available globally since they are a
major fruit in many countries.
The cheap source of anthocyanin is by-products of some industrial practices like
grape skin extracts. Therefore more focus is being given to anthocyanin-rich waste
by-products like purple corncobs and banana bracts. The use of these by-products is
however very challenging. Other ingredients can be added. Designing an accurate
recipe of different reagents to increase their color stability as well suitable process-
ing and storage time can lead to stable colors.
Although being obtained from plants, there are many challenges. For example
the supply is subjected to long cultivation times, climatic and seasonal variations,
disease risks, increasing cost of agriculture as well as declining availability of culti-
vable land all making their availability highly challenging. New feasible candidate
to be investigated as new natural pigment must be readily available, economical and
high yield plant and with satisfactory tinctorial strength. The great activity in this
fi eld is supported by data from a survey by Frost and Sullivan in 2002, who predict
an expansion of the European color market by 1 % per year till 2008, whereas color-
ing foodstuff is estimated to grow even by 10–15 % in the same time range [ 12 ].
4.2 Use of Anthocyanin-Based Colorants
Anthocyanins are being used by humans since beginning of civilization for decora-
tive purposes. Egyptians used wine as source of food color to enhance the color of
candies approximately 1500 BC. The oldest anthocyanin used and marketed is enocy-
nain obtained from pomace of red grape in Italy since 1879. According to the USA
regulations, “color additives are defi ned as any dye, pigment or substance capable of
imparting color when added or applied to a food, drug or cosmetic (21CFR70.3). ANs
are permitted as food colorants in the USA under the category of fruit (21CFR73.250)
or vegetable (21CFR73.260) juice color.” Under these categories only suitable for
eating part of plants and their water extracts are permitted for use.
Anthocyanins themselves and any of anthocyanin-derived colorant are recog-
nized as a natural colorant (labeled as E163) by the European Union in Codex
Alimentarius system. In USA, the Food and Drug Administration Authority (FDA)
has cataloged anthocyanin under list of natural colors that do not require certifi ca-
tions, i.e. without FD&C numbers. Out of these exempted 26 colorants approved by
4.2 Use of Anthocyanin-Based Colorants
52
FDA, 4 are based on anthocyanin pigments. Anthocyanins are a potent alternative to
FD&C Red No. 40 which is a synthetic dye with highest consumption in US. Various
food plants have been considered as marketable source of anthocyanin-based colo-
rants, however their use has been limited by pigment stability, availability of raw
material and economic considerations [ 3 ].
4.3 Acylated Anthocyanins as Colorants
for the Food Industry
Anthocyanins are potential food color substitute however the major demerit is poor
stability, however acylation perk up color and pigment stability. These acylated
anthocyanins have variety of shades that are dependent on pH of food matrix and
structure changes in anthocyanins. For example acylated pelargonidin derivatives
obtained from radish and potato at acidic pH match allura red and acylated cyanidin
from black carrots or cabbage pH dependent hues ranging from deep red to purple.
More than 65 % of reported anthocyanins whose structures have been properly
identifi ed are acylated. Anthocyanins diversity greatly depends on number, nature
and linkage position of acyl groups.
Acyalted derivatives of anthocyanins are stable and have wide color range so
scientists are in search of acylated anthocyanin-rich material (Table 4.1 ) for exam-
ple red cabbage in Polish food industry [ 13 , 14 ] has color like blueberries at pH 4.2–
4.5 and usually the extract of red cabbage is available in a liquid or sprayed-dried
water-soluble form [ 3 ]. Another exceptional source of acylated anthocyanins is
black carrot ( Daucus carota L) consumed in Afghanistan, Egypt, Pakistan, Spain,
and Turkey [ 3 ]. It is well-known now that poly acylated anthocyanins are more
stable while simple anthocyanins quickly drop their colors by hydration at 2- position
of anthocyanidin nucleus.
Another acceptable source of stable acylated anthocyanins is red potatoes [ 15 ,
16 ]. Extracts of acylated anthocyanins can be applied for foods having a low pH
level, including jellies, sauces, conserves, confectionery and bakery items and soft
drinks [ 17 ]. Some other potential applications of acylated anthocyanins are neutral
or slightly alkaline products, such as powdered and ready-to-eat desserts, milk-
drinks, ice cream and panned products [ 3 , 17 ]. Table 4.1 shows sources of some
acylated anthocyanins, their properties and their use in food industry.
Anthocyanins react very rapidly with metals forming stable complexes with iron,
copper and tin, these complexes can be used as potential coloring agents due to their
higher stability. Acylation decrease water solubility while glycosylation increases
it. Hydroxylation (with more –OH groups) produces blue colors while methoxyl-
ation (with more methoxyl groups) produce red colors. The number and type of
glycosidic units at C-3 don’t change color signifi cantly while additional glycosyl-
ation at C-5 induces a slight shift to red-purple.
4 Anthocyanins as Natural Colors

53
Table 4.1 Sources of acylated anthocyanins, their properties and their uses in food industry
Source Properties Types of anthocyanins Trade name Company
Maqui berry ( Aristotelia
chilensis L)
Excellent ROS/RNS scavenger, ↓
oxidative stress, blood glucoe, platelet
activity
Derivatives of delphinidin
3,5-O-diglucoside, delphinidin
3-O-samb, 5-O-glucoside, delphinidin
3-O-glucoside
Delphinol
®
Maqui New Life
Red cabbage ( Brassica
oleracea L)
Expensive, produces different shades at
different pH, have pleasant taste,
stability to heat and light, low in
polyphenols reducing the risk of hazing
with proteins, available all year round
and have potential health benefi ts when
included into the diet
15 Anthocyanins (mainly diacylated)
being derivatives of cyanidin-3-
diglucoside, -5-glucoside acylated with
sinapic, ferulic and/or p -coumaric acids
Magento™ Overseal Foods Ltd
Black carrot ( Daucus
carota L)
Provide an excellent bright strawberry
red shade in acidic products, exhibit
mauve to blue tones under neutral pH,
vegetarian and kosher alternative to
carmine, has low levels of polyphenols
and improved stability to heat, light and
SO
2
Derivatives of cyanidin-3- rutynoside-
-glucoside- galactoside acylated with one
cinnamic acid ( p - -coumaric, ferulic or
sinapic)
Exberry
®
Carantho
®
GNT Group,
Overseal Foods Ltd
Red radish ( Raphanus
sativus L).
Acylated pelargonidin derivatives from
red radish give red color to maraschino
cherries very close to that of synthetic
colorant FD&C Red # 40 at pH 3.5
This vegetable contains 12 acylated
anthocyanins (8 diacylated)
ColorPure™ RFI
Red potatoes ( Solanum
tuberosum )
Possible natural alternative pigment
source to synthetic colorant FD&C Red
# 40
Monoacylated anthocyanins Not available Nil
Purple corn ( Zea mays )
and oxalis ( Oxalis
triangularis )
Under consideration as potential food
color sources
Not yet known Not available Nil
4.3 Acylated Anthocyanins as Colorants for the Food Industry
54
4.4 Conclusions
Pigments act as fi ngerprint of a food commodity determining its overall quality.
Anthocyanin-rich extracts obtained from purple corn, blackcurrant, grape skin and
red cabbage are now approved as natural colorants in New Zealand, Australia and
Europe and are commercially used in food industry. However, anthocyanins as nat-
ural colors have their own limitations. Diffi culties in collection of plant materials,
lack of standardization, extraction procedure from plants material and low color
value makes them less focused as substituent to synthetic colors. If somehow
obtained and used, they face problem of instability due to temperature, oxygen,
light, pH and enzymes. Further the color imparted is fugitive sometimes and a mor-
dant needs to be added to increase colors. However it should not be a disappointing.
Their stability can be increased by adding stabilizers like dextrin additives obtained
from tart cherries. Further research should be conducted for identifi cation, charac-
terization, documentation, assessment and potential utilization from un-prospected
plant species. The large scale cultivation from these species and increasing anthocy-
anin contents in existing species by genetic engineering will lead to availability of
suffi cient amounts for industrial extraction, improvement of quality and quantity of
anthocyanins and enhance their stability during processing and storage.
References
1. Delgado-Vargas, F., Jiménez, A., & Paredes-López, O. (2000). Natural pigments: Carotenoids,
anthocyanins, and betalains—Characteristics, biosynthesis, processing, and stability. Critical
Reviews in Food Science and Nutrition, 40 (3), 173–289.
2. Current EU approved additives and their E numbers. (2014). EU Food Standard Agency .
http://
www.food.gov.uk/science/additives/enumberlist
.
3. Giusti, M. M., & Wrolstad, R. E. (2003). Acylated anthocyanins from edible sources and their
applications in food systems. Biochemical Engineering Journal, 14 (3), 217–225.
4. Mazza, G., & Miniati, E. (1993). Anthocyanins in fruits, vegetables, and grains . Boca Raton:
CRC.
5. Harborne, J. B., & Williams, C. A. (2001). Anthocyanins and other fl avonoids. Natural
Product Reports, 18 (3), 310–333.
6. Kong, J. M., Chia, L. S., Goh, N. K., Chia, T. F., & Brouillard, R. (2003). Analysis and biologi-
cal activities of anthocyanins. Phytochemistry, 64 (5), 923–933.
7. Strack, D., Steglich, W., & Wray, V. (1993). Betalains. Methods in Plant Biochemistry, 8 ,
421–450.
8. Pietta, P.-G. (2000). Flavonoids as antioxidants. Journal of Natural Products, 63 (7),
1035–1042.
9. Andersen, Ø. M., Fossen, T., Torskangerpoll, K., Fossen, A., & Hauge, U. (2004). Anthocyanin
from strawberry ( Fragaria ananassa ) with the novel aglycone, 5-carboxypyranopelargonidin.
Phytochemistry, 65 (4), 405–410.
10. Wrolstad, R. E., Durst, R. W., & Lee, J. (2005). Tracking color and pigment changes in antho-
cyanin products. Trends in Food Science and Technology, 16 (9), 423–428.
4 Anthocyanins as Natural Colors
55
11. Anderson, Ø., & Jordheim, M. (2008). Anthocyanins: Food applications . In Proceedings of 5th
International Congress Pigments in Food: For Quality and Health, University Helsinki
Helsinki, Finland .
12. Stintzing, F. C., & Carle, R. (2004). Functional properties of anthocyanins and betalains in
plants, food, and in human nutrition. Trends in Food Science and Technology, 15 (1), 19–38.
13. Baublis, A., Spomer, A., & Berber Jiménez, M. D. (1994). Anthocyanin pigments: Comparison
of extract stability. Journal of Food Science, 59 (6), 1219–1221.
14. Giusti, M. M., Luis, E. R., Donald, G., & Ronald, E. W. (1999). Electrospray and tandem mass
spectroscopy as tools for anthocyanin characterization. Journal of Agricultural and Food
Chemistry, 47 (11), 4657–4664.
15. Fossen, T., & Andersen, Ø. M. (2003). Anthocyanins from red onion, (Allium cepa) with novel
aglycone. Phytochemistry, 62 (8), 1217–1220.
16. Terahara, N., Konczak, I., Ono, H., Yoshimoto, M., & Yamakawa, O. (2004). Characterization
of acylated anthocyanins in callus induced from storage root of purple-fl eshed sweet potato,
Ipomoea batatas L. BioMed Research International, 2004 (5), 279–286.
17. Naturex . (2014). Ultimate botanical benifi ts . Cited September 2014.
References
57
© The Author(s) 2016
M. Riaz et al., Anthocyanins and Human Health, SpringerBriefs in Food,
Health, and Nutrition, DOI 10.1007/978-3-319-26456-1_5
Chapter 5
Anthocyanins Absorption and Metabolism
5.1 Introduction
Biologically-active compounds, after ingestion, interact during metabolism to mod-
ify biological responses that can either strengthen or restrict their effi cacy and
potency. Regarding anthocyanins, as discussed before in Chaps. 2 and 3 , no member
of animal kingdom can synthesize these molecules and they have to rely on plants.
Although humans are not colored by anthocyanins, low to signifi cant contents of
anthocyanins are observed in human blood, tissues and organs. Demographical
studies indicate that most of the world population lives in poverty and do not have
luxury to afford such dietary recommendations. The effi ciency of food in digestion
and bioavailability is the chief factor, i.e. the amount of anthocyanins released, solu-
bilized, absorbed, transported and metabolized. It is once the anthocyanins are in
human body that the major uncertainty begins. The ratio of total digested anthocya-
nins to bioavailable anthocyanins must always be considered. The outcomes are
highly inconsistent and sometimes contradictory. Due to variations of experimental-
designs, analytical procedures and species analyzed, the statistical fi gures reported
and arithmetical comparisons between studies are not precise.
Stability and functionality of anthocyanins in human body depends on their con-
tent, type, and location in food matrix and presence of other bioactive compounds in
fruits and vegetables. Their instability depends directly on number of –OH groups
and indirectly on number of –OCH
3
groups. Further glycosylation also affects stabil-
ity as diglucoides are more stable than monoglucosides. Being polar in nature, antho-
cyanins are more soluble in polar solvents as compared to non-polar solvents. Further
food processing can lead to qualitative and quantitative changes since anthocyanins
are susceptible to oxidation and isomerization. Their bioeffi cacy depends upon their
bioavailability and bioaccessibility. Studies indicate that bioavailability is variable
depending upon various diverse parameters. Anthocyanidins and anthocyanins are
the least absorbed phenolics. Therefore, their abundance in term of dietary intake
does not necessarily mean their intact and relative metabolites in human body.
58
The main criterion for absorption and resultant exerted bioactivity is liberation
from food matrix, i.e. bioaccessibility. Bioaccessibility indicates the amount of
anthocyanins available for absorption in gut. Bioaccessibility and bioavailability
depends on the physicochemical characteristics of food matrix which affects the
effi cacy of the enzymatic and physicochemical digestion processes. Actually food
micro-structure determines release, transformation and absorption of anthocyanins
in digestive tract. Complexation with other food components, attachment to specifi c
organelles and restriction within intact cell walls or entrenchment in food matrix
may affect the release of anthocyanins. Consequently anthocyanins bioavailability
and bioactivity depends upon food matrix behavior, the presence of promoters and
inhibitors of anthocyanins absorption. Food processing disrupts the food matrix
which can affect their bioavailability. While the analytical methods themselves are
subject to various sources of errors, the main problem with anthocyanins analysis
lies in their inherent instability.
5.2 Daily Intake
Currently, we are watching a worldwide sharp increase in research activity as well
as in dietary supplement containing anthocyanins. Despite the fact that the need for
anthocyanins is different and depends on income level, information of anthocyanins
content and composition is necessary so that guidelines can be provided on food
sources that can provide ample to rich supplies of preferred anthocyanins.
Anthocyanins are quickly being incorporated into the mainstream channel of func-
tional foods, dietary supplements and nutraceuticals and progressively being recog-
nized by the public due to the demand for health-promoting diets. Depending upon
the dietary habits and socioeconomic conditions, the daily intake of anthocyanins
varies from micrograms to hundreds of grams per person. Although individuals in
developed countries use more anthocyanins due to awareness of their health-
promoting effects created by television commercials and all types of media-
campaigns, intake of diets containing anthocyanins is gradually increasing in
low-income countries also because of easy commercial and market availability of
juices, drinks, slushes, concentrates and extracts of fruits and vegetables containing
greater anthocyanins contents. Since anthocyanins are not classifi ed as essential
nutrients so their recommended daily intake (RDI) has not been established yet.
Daily intake of anthocyanins in USA had previously (1976) been estimated to be
180–215 mg per day per person [ 1 ], but a recent study by USDA, based on the
evaluation of more than 100 common foods in the USA, daily intake was found to
be 12.5 mg per day per person [ 2 ]. Interestingly 100 g of berries offer up to 500 mg
of anthocyanins [ 3 ]. This intake of anthocyanins is higher compared with dietary
fl avonoids such as genistein and quercetin (estimated at 20–25 mg/day) [ 4 ]. The
daily intake of anthocyaninsis higher compared than in the USA. According to mul-
tinational study across ten European countries report (36,037 individuals in the age
of 35–74 years), the mean daily intake of anthocyanidins [ 5 ] ranged in men from
5 Anthocyanins Absorption and Metabolism
59
19.8 to 64.9 mg, while in women from 18.7 to 44.1 mg. In addition, this study
showed high anthocyanidins intake by southern European, non-obese and non-
smoker older women [ 6 ]. The different values of daily intake are mainly due to
different food intake data. Authentic and exact food intake data along with complete
food anthocyanin concentration data is necessary for estimation of daily intake.
Proanthocyanidins content of food fl uctuates depending upon distribution of
monomers (fl avan-3-ols), oligomers and polymers (usually N10, fl avan-3-ol mono-
mers), and their inter-fl avan bonds [ 7 ]. The mean daily intake of oligomeric and
polymeric proanthocyanidins was estimated to be higher than that of monomeric
fl avan-3-ols, and twice as high as the combined overall intake of other fl avonoids
[ 8 ]. The total proanthocyanidin intake by American adults range from 96 to 137 mg
per day (mg/day) [ 9 ],whereas those reported for Spanish 189 mg/day and for Finnish
adults was [ 10 ] 128 mg/day [ 11 ]. Wang et al. (2011) recorded the mean daily intake
in American adults to be 95 mg/day [ 12 ], the value of intake lies at lower end of
amount that was previously seen by Gu et al. [ 9 , 13 ].
5.3 Anthocyanins Absorption
The preventive and therapeutic actions of anthocyanins are directly proportional to
their absorption and metabolism in human body. Many diverse challenges exist in
conducting kinetic studies in humans mainly due to their low levels in circulation.
Comprehensive information of qualitative and quantitative distribution of anthocya-
nins in food chain, human blood, tissues and organs is crucial in order to establish a
statistically signifi cant correlation between consumption of anthocyanins and their
preventive and pharmacological effects. The absorption of anthocyanins depends
upon many factors including chemical structure, nature of food matrices, interaction
with other micro, macro and phytonutrients, type and extent of food processing and
preparation, as well as the nutritional, pathophysiological and genetic factors of
individual. The change in absorption of anthocyanins is also associated with its
biological action, in general the absorption of the anthocyanins are less, so this point
may be used for increasing the biological actions by playing with anthocyanins
pharmacokinitics via different ways. However the structural and functional differ-
ences should be considered with respect to the source of anthocyanins for example
the vegetable anthocyanins are more complex than fruit and thus their hydrolysis
rate, bioavailability and metabolism is different.
Various factors can infl uence the ability to absorb, convert and metabolize the
anthocyanins. Due to their nutritive and health promoting effects, interest in factors
that infl uence their bioavailability has intensifi ed. Biological activities of anthocya-
nins are dependent on its bioavailability, i.e. the amount of anthocyanins reach to
systemic circulation and it is usually based on absorption and metabolism. Despite
much information, still our knowledge about anthocyanins distribution and
availability is limited. Usually acylated anthocyanins are less absorbed than non-
acylated ones. The type and numbers of backbone and acyl group are believed to
5.3 Anthocyanins Absorption
60
affect their bioavailability in humans. Bioavailability from supplements is usually
much better. In mouth, chewing releases anthocyanins from the food matrix. The
bioavailability of anthocyanins is very low, with <1 % of the ingested amount reach-
ing the plasma, glucuronidated and methylated anthocyanin metabolites were found
twice than their parent (intact) compounds [ 14 ]. In human body anthocyanins inter-
act physically and chemically with other phytochemicals which may lead to syner-
gism or interference.
5.3.1 Gastric Absorption
Anthocyanins absorption may start in the stomach and appear in the blood immedi-
ately after ingestion. Passamonti et al. (2003) and Talavera et al. (2003) used analo-
gous methods to establish that anthocyanins were profi ciently absorbed in the
stomach [ 15 , 16 ]. Passamonti et al. injected grape anthocyanins into the stomach of
19 Wistar male rats and collected blood from both the portal vein and the heart at
6 min. Malvidin-3-glucoside was present in both portal and systemic plasma.
Importantly, malvidin-3-glucoside appeared in the plasma within 6 min, presenting
an evidence of stomach absorption. Peonidin-3-glucoside, petunidin-3-glucoside,
and malvidin-3-acetyl glucoside derivatives were inconsistently detected, perhaps
owing to animal variability. Neither anthocyanins aglycones nor conjugated deriva-
tives were detected in the plasma. Talavera et al. [ 16 ] (2003) infused anthocyanin
standards as well as bilberry and blackberry extract into the stomach of pylorus- and
sphincter-ligated rats. Gastric contents sample from gastric vein and blood sample
from abdominal aorta were taken 30 min after the administration. HPLC analysis
revealed that a high proportion (∼25 %) of anthocyanin mono-glycosides, including
glucoside and galactoside, was absorbed from the stomach, whereas the rutinoside
was poorly absorbed. It was suggested that gastric absorption of anthocyanins
involves bilitranslocase (TC 2.A.65.1.1), an organic anion membrane carrier in the
gastric mucosa [ 17 ].
5.3.2 Absorption in the Small Intestine
The small intestine is the major site for anthocyanins absorption. Absorption of
anthocyanins in small intestine of anesthetized rats was estimated by an in situ per-
fusion method [ 18 ] supplemented in physiological buffers. The amount of antho-
cyanins remaining in the effl uent was used to estimate the rate of anthocyanins
absorption in the small intestine. Depending on their structures, the absorption rate
of supplemented anthocyanins ranged from 22.4 ± 2.0 % (cyanidin-3-glucoside) to
10.7 ± 1.1 % (malvidin-3-glucoside). Such high absorption rates seemed to contra-
dict the very low levels of anthocyanins observed in the blood [
19 ]. However, it has
to be noted that these absorption rates were calculated based on the disappeared
5 Anthocyanins Absorption and Metabolism
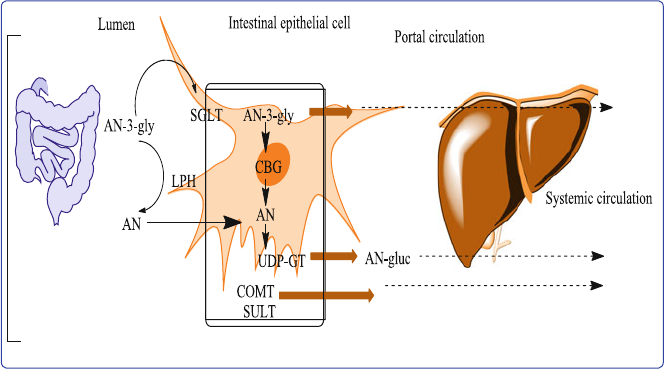
61
amount in the effl uent, thus they could indicate the portion of anthocyanins being
taken up into the small intestine tissue but not necessarily transferred into the blood.
Recently, our research group also demonstrated that as high as 7.5 % of the admin-
istered black raspberry anthocyanins could be taken up by rat small intestinal tissue,
yet very limited amount can be detected in urine [ 20 ].
Endogenic β-glucosidases are involved at this stage to release aglycones from
anthocyadin-glucosides. Since anthocyanin are highly water soluble and are very
large molecules, their absorption cannot take place by passive diffusion. Therefore
absorption of anthocyanins should be either hydrolyzed to the aglycone in small
intestine by α-rhamnosidase, β-glucuronidase or β-glucosidase or they should uti-
lize an active transport mechanism to transport glycosides across the intestinal wall.
Aglycones in free status are more hydrophobic and are smaller than the glycosides
and can easily infi ltrate the epithelial layer passively. However intact glycosides are
absorbed by the small intestine, either by sodium-dependent glucose transporter
(SGLT1) or by the ineffi cient passive diffusion. Recent evidence have suggested
that acylated anthocyanins are somewhat bioavailable in the intact form [ 21 ],
although, likely owing to their increased molecular size, acylated ANs are much
less effi ciently absorbed than their counterparts without the acylation [ 22 ].
Phase II enzymes convert anthocyanins to glucuronids, methylates and sulfates
in liver and kidney [ 23 , 24 ]. These conjugated forms of anthocyanins may be
excreted via bile to jejunum and recycled by enterohepatic circulation system in
intestine/colon. Figure 5.1 indicates the potential mechanism of anthocyanins (ANs)
absorption
Fig. 5.1 Potential mechanism of anthocyanins (ANs) absorption, sodium-glucose co-transporter,
AN-3-gly anthocyanins-3-glycoside, CBG cystolic B-glucosidase, LPH lactate pholorizin hydro-
lase, UDP-GT UDP-glucuronosyl-transferase, AN-gluc anthocyanins glucuronide, COMT
catechol- O-methyl = transferase, SULT sulfotransferase [
25 ]
5.3 Anthocyanins Absorption
62
Many studies have confi rmed the blood circulation of anthocyanins after fast-
track absorption in intact form from stomach and small intestine and their excretion
as methylated, sulfo-conjugated or glucueonide derivatives via urinary system [ 15 ,
16 , 23 , 26 ]. These are one of few anthocyanins that are found in intact forms in
blood plasma [ 27 ]. In a recent study when anthocynain-rich diet was fed to rats for
almost 17 days, their presence was observed in stomach, small intestine, liver, kid-
ney and brain. Total anthocyanin content in brain (blackberry anthocyanins and
peonidin 3- O -glucoside) was found 0.25 ± 0.05 nmol/g of tissue [ 14 , 28 ].
5.3.3 Pharmacokinetics
The knowledge of pharmacokinetics is vital to comprehend the effect of daily intake
of anthocyanins on health-improvement. There is limited information on complete
pharmacokinetic data of anthocyanins. Challenges exist in understanding kinetics
of anthocyanins in biological systems due to their low levels in human circulation
making it hard to observe their absorption and metabolism. Despite this limitation,
considerable work has been carried out in last decade in assessing the dynamics and
kinetics of anthocyanins in humans. Figure 5.2 shows an overview of anthocyanins
(ANs) absorption and excretion.
Previous studies indicate that average quantity of anthocyanins in blood plasma
vary from 1.4 and 592 nmol/l followed by post-consumption of dose from 68 and
1300 mg between 0.5–4 h. Further, the average urinary excretion of anthocyanins
varies from 0.03 and 4 % of the total ingested anthocyanins, and maximum excre-
tion occurs between 1–4 h and having a t
1/2
of 1.5–3 h. Although quercetin are
excreted slowly (t
1/2
11–28 h), which indicates the probability of their bioaccumula-
tion [ 29 ], the excretion of anthocyanins is pretty quick (t
1/2
1.5–3 h) and no signifi -
cant bio-accumulation occurs after a diet containing normal amount of anthocyanins.
However the importance of increase of colonic metabolites and aglycone metabolic
products should be explored.
Pharmacokinetic investigations indicate the presence of anthocyanins and their
glucuronide derivatives in blood up to 5 h after intake while transformation into
methyl derivatives increases over time (6–24 h). Another study also confi rmed the
presence of metabolites with their basic anthocyanin skeleton in the urine for up to
24 h [ 26 ] thus biological activity changes with metabolic transformation e.g. glyco-
sylation and acylation patterns decrease the bioavailability of an anthocyanin; how-
ever, glycosidases present in the GIT may hydrolyze anthocyanins into
anthocyanidins, thereby increasing their biological potential but decreasing their
stability. The presence of a glucose moiety compared with a galactose or arabinose
on the cyanidin and peonidin anthocyanidins present in cranberry juice seems to
make them more bioavailable as a percentage of the delivered dose [ 30 ].
The absorption of anthocyanins from food is insuffi cient and the contents detected
in plasma are from nM to low μM [ 26 , 28 ]. Kay et al. [ 26 ], reported the total cumula-
tive concentration 172.96 ± 7.44 μg h/mL of anthocyanins (parent and metabolites)
5 Anthocyanins Absorption and Metabolism
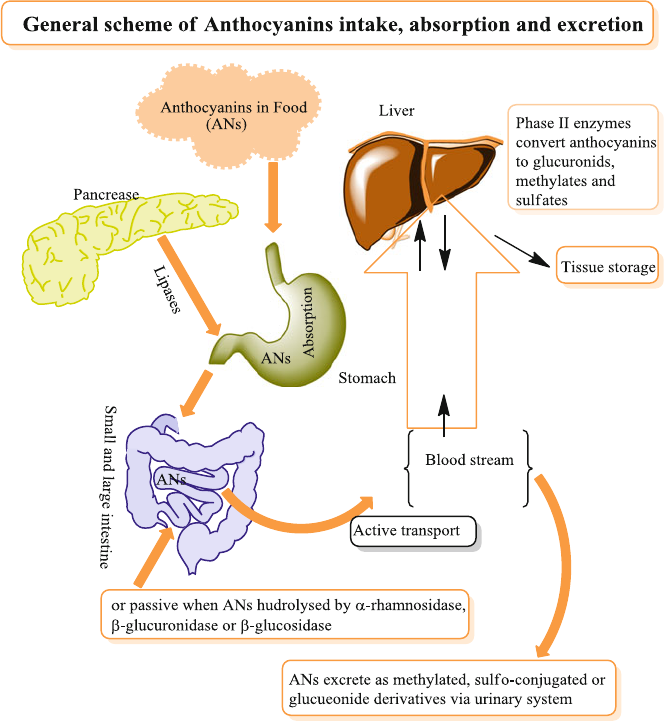
63
detected in the serum over a 7 h sampling, peak plasma level 44.86 ± 2.82 μg/mL
was achieved (C
max
) within 2.8 h (t
max
), cyanidin 3-glycosides was found the parent
of the total anthocyanins in the serum. Further, the total urinary excretion of metabo-
lites and parent compounds over 24 h was 1071.54 ± 375.46 μg, reaching a maximal
rate of excretion (R
max
) of 202.74 ± 85.06 μg/h at 3.72 ± 0.83 h (t
max
) and having an
elimination half-life (t
1/2
) of 4.12 ± 0.4 h. Correspondingly, only 32.5 % of the antho-
cyanins excreted in the urine were the parent compounds with an average of 67.5 %
occurring as conjugated metabolites.
To exert any physiological effect, bioactive molecules should be bioavailable.
The percentage of anthocyanins, both native and metabolites, absorbed and evacu-
ated in urine is extremely small in-comparison to consumed amount. The bioavail-
ability of anthocyanins is very low, with <1 % of the ingested amount reaching the
Fig. 5.2 An overview of anthocyanins (ANs) absorption and excretion
5.3 Anthocyanins Absorption
64
plasma, glucuronited and methylated anthocyanin metabolites were found twice
than their parent (intact) compounds [ 14 ].
The reason for low bioavailability of anthocyanins can be underestimation that
may occur because concentration of some metabolites, such as protocatechuic acid,
might be below the detection limit of the analytical methods employed and the pre-
dominance of the colorless carbinol (75–80 %) and chalcone (15–20 %) forms of
anthocyanins present in blood and urine at neutral pHs, it is highly likely that these
chemical forms may escape detection, and/or be chemically bound to other compo-
nents in the plasma or urine, and therefore not included in the analyzed fraction.
Labeling anthocyanins for identifi cation of all generated metabolites may overcome
these shortcomings [ 14 , 27 ].
5.4 Carbohydrates Moieties Deconjugation
Glycosylated anthocynains are better hydrophilic as compared to aglycosylated
(aglycones) ones and passively diffuse the cell membrane due to their more lipid
solubility and small molecular size. During this passage, ingested anthocyanins are
deglycosylated. Despite the strong acidic conditions of stomach, there are very little
chances of acid hydrolysis (non-enzymatic deglycosylation). Deglycosylation of
glycosylated anthocyanins [ 31 ] was not observed after pepsin-HCl digestion even at
stronger conditions (pH 2.0, 37 °C, 2 h). There are chances of in vivo enzymatic
deglycosylation as observed by using animal models although very little evidence is
available to support this path. GIT of rats and pigs showed selective degradation of
anthocyanin-glucosides in small intestine [ 2 , 20 ], however further characterization
of deglycosylation patterns of anthocyanins under the effect of isolated small intes-
tinal β-glucosidases is needed. Even in rat small intestine in situ perfusion model,
the disappearance of cynadin-3-glucoside was signifi cantly higher than other glyco-
sides of cynadin [ 18 ]. Limited information available suggests that anthocyanins
containing xylose are better retained in the fecal content and feces as opposed to
anthocyanins containing galactose and glucose [ 21 ]. Detailed studies are required to
interpret the destiny of such glycosides.
Various factors infl uence the bioavailability of anthocyanins, like quantity of gut
microfl ora, food matrix [ 32 ], analytical detection problems (e.g. carbinol and chal-
cone) and differences in xenobiotic metabolism in GIT, liver, and other tissues.
5.5 The Infl uence of Colonic Microfl ora
Gut microfl ora converts anthocyanins into less stable anthocyanidins at neutral pH
leading to degradation within 20 min thus Cyanidin-3-rutinoside was fi rst hydro-
lyzed into cyanidin-3-glucoside and then into cyaniding aglycon, which rapidly
degraded into protocatechuic acid (3,4-dihydroxybenzoic acid) [ 33 ]. Some
5 Anthocyanins Absorption and Metabolism
65
anthocyanins metabolites have greater microbial and chemical stability, indicating
their potential role in various biological effects and enhancement of antioxidant
activity [ 33 ]. It is pertinent to mention that degradation of methylated anthocynains
by the gut microbiota may yield de-methylated products [ 27 ].
Since there are no endogenous esterases in humans to release phenolic acids,
therefore the esterase activity of colonic microfl ora is required for the metabolism of
acylated fl avonoids [ 34 ]. Aura et al. [ 35 ] revealed that human fecal fl ora immediately
deconjugates quercitin glucosides, -rutinoside and glucuronide (glc). Simple pheno-
lics like 3,4-dihydroxyphenyl acetic acid and its derivatives are generated during ring
fi ssion of deglycosylated quercetin [ 36 ]. Aura et al. suggested that gut microfl ora can
convert anthocyanins as revealed by fermentation of cyanidin-3- rutinoside and cyan-
idin-3-glucoside in the presence of human fecal slurry. Cyanidin- 3- glucoside was
nearly hydrolyzed entirely after incubation of 2 h, and smaller than one third of the
cyanidin-3-rutinoside present. Protocatechuic acid (PC), a ring- fi ssion product of
cyanidin-aglycone, was the main metabolite [ 35 ]. More than 90 % of the cyanidin-3,
5-glucoside after incubation with human fecal suspension was degenerated after 2 h
and fractional hydrolysis yielded cyanidin mono- glucoside as an intermediate of
deterioration, which also degraded in the meantime. Resultant production and piling-
up of protocatechuic acid was observed again. Assessment of di-acylated anthocya-
nins from red radish indicated that the acyl group can be broken down by fecal
microfl ora and the released acids were comparatively stable [ 37 ]. Deacylated antho-
cyanins can follow the same pathway of degradation as discussed above.
5.6 Metabolism in Intestinal Mucosa and Tissues
The polyphenolic and cationic attributes of anthocyanins and their metabolites acti-
vate a variety of cellular responses. Some metabolites are responsible for bioactivity
of fl avonoids. For example, methylated cyanidin-3-glucoside is transformed to
peonidin-3-glucoside [ 38 ]. Similarly, certain metabolic products of anthocyanins
may have superior activity as compared to parent molecules.
5.7 Tissue Distribution
The knowledge of tissue distribution of anthocyanins is necessary to evaluate the
health-promoting effects of anthocyanins. This information will also help to screen
the suitable anthocyanins from various sources to assist development of supple-
ments and functional foods. The protective potential of anthocyanins associated
with various tissue-diseases is proved in vitro by using various types of cell lines.
However in vivo studies of same are limited. Distribution of anthocyanin in tissues
has recently been evaluated in pig and rat models. Male Wistar rats were fed black-
berry extract (370 nmol anthocyanin/day) for 15 days and killed at 3 h after the
5.7 Tissue Distribution
66
beginning of the last meal. Total anthocyanins averaged 605 nmol g
−1
in jejunum,
68.6 nmol g
−1
in stomach, 3.27 nmol g
−1
in kidney, 0.38 nmol g
−1
in liver, and
0.25 nmol g
−1
in brain [ 28 ]. In pigs fed diets supplemented with 0, 1, 2, or 4 % w/w
blueberries for 4 weeks and fasted for 18–21 h before euthanasia, 1.30 pmol g of
anthocyanins were identifi ed in the liver, 1.58 pmol g
−1
in eyes, 0.878 pmol g
−1
in
cortex, and 0.664 pmol g
−1
in cerebellum [ 39 ]. The results suggested that anthocya-
nins after crossing the blood-retinal and the blood-brain barrier can protect brain
and eye tissues respectively. This hypothesis is also favored by another research in
which aged blueberry-fed rats were used [ 40 ].
5.8 Excretion
Urine is the major excretory pathway of intact anthocyanins and aglycones [ 41 ].
Although most conjugated fl avonoid metabolites follow the same route [ 38 ], how-
ever a small portion of them may re-enter the jejunum via bile, and are absorbed by
the colon entering the entero-hepatic circulation again [ 42 ], or are defecated with
feces. The lung is a chief excretion place for many bioactive constituents including
quercetin [ 43 ]. However, there is no evidence indicating excretion of anthocyanin
by respiration.
Recently biodegradation of anthocyanins into phenolic acids by enteric bacteria
has gained much attention (Table 5.1 ). Various studies have documented the pres-
ence of phenolic acids, like protocatechuic acid, gallic acid, vanillic acid, phloroglu-
cinol acid, syringic acid and phloroglucinol aldehyde anthocyanin metabolites
[ 45 – 47 ]. These phenolic acids are produced during anthocyanin metabolism either
by enteric bacteria, or by chemical conversion, and have also been detected in
humans [ 48 ].
Anthocyanidin aglycones are degraded much faster as compared to anthocyanins
monoglycosides [ 33 ]. Similarly anthocyanins degradation by intestinal microfl ora
is also faster [ 44 ]. Unabsorbed and non-degraded anthocyanins are excreted in their
intact forms. Unchanged anthocyanins are detected in fecal samples of humans 24 h
after ingestion of blood orange juice [ 49 ], and in fecal samples of rats fed by grapes,
bilberries and chokeberries [ 21 ].
Table 5.1 Degradation
products of anthocyanidin
aglycons [
44 ]
Anthocyanidin
aglycon
Corresponding
phenolic acid
Cyanidin → Protocatechuic acid
Delphinidin → Gallic acid
Pelargonidin → 4-Hydroxybenzoic acid
Malvidin → Syringic acid
Peonidin → Vanilic acid
Petunidin → 3-O-methylgallic acid
5 Anthocyanins Absorption and Metabolism
67
5.9 Conclusions
The benefi cial effects of anthocyanins have made the study of their bioavailability,
absorption and metabolism an area of considerable interest since humans can’t syn-
thesize these vital molecules de novo and obtain them from plant kingdom.
Understanding of anthocyanin chemistry, metabolism and their mechanism of
action has increased substantially in previous 10 years. Many older questions and
controversies have been resolved. Studies have been conducted in last decade to
develop a more thorough understanding of factors that infl uence their uptake,
metabolism, and utilization in humans. This information however is only tip of the
iceberg. The exact molecular mechanism behind their metabolism remains to be
fully established. Although not all animals are good models for metabolic studies of
anthocyanins, rats are commonly used. There is need for validation of new animal
models especially induced mouse mutants in studying anthocyanin metabolism.
The effects of variety, growing practices, season, stage of maturity, storage and
processing condition are important for researchers to understand anthocyanin
metabolism in epidemiological studies.
References
1. Kühnau, J. (1976). The fl avonoids. A class of semi-essential food components: Their role in
human nutrition. World Review of Nutrition and Dietetics, 24 , 117.
2. Wu, X., Pittman, H. E., & Prior, R. L. (2006). Fate of anthocyanins and antioxidant capacity in
contents of the gastrointestinal tract of weanling pigs following black raspberry consumption.
Journal of Agricultural and Food Chemistry, 54 (2), 583–589.
3. Mazza, G., & Miniati, E. (1993). Anthocyanins in fruits, vegetables, and grains . Boca Raton:
CRC.
4. Hertog, M. G., Feskens, E. J., Hollman, P. C., Katan, M. B., & Kromhout, D. (1993). Dietary
antioxidant fl avonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet,
342 (8878), 1007–1011.
5. Zamora-Ros, R., Andres-Lacueva, C., Lamuela-Raventós, R. M., Berenguer, T., Jakszyn, P.,
Martínez, C., et al. (2008). Concentrations of resveratrol and derivatives in foods and estima-
tion of dietary intake in a Spanish population: European Prospective Investigation into Cancer
and Nutrition (EPIC)-Spain cohort. British Journal of Nutrition, 100 (01), 188–196.
6. Tsuda, T. (2012). Anthocyanins as functional food factors-chemistry, nutrition and health pro-
motion. Food Science and Technology Research, 18 (3), 315–324.
7. Prior, R. L., & Gu, L. (2005). Occurrence and biological signifi cance of proanthocyanidins in
the American diet. Phytochemistry, 66 (18), 2264–2280.
8. Hellström, J. K., Torronen, A. R., & Mattila, P. H. (2009). Proanthocyanidins in common food
products of plant origin. Journal of Agricultural and Food Chemistry, 57 (17), 7899–7906.
9. Gu, L., Kelm, M. A., Hammerstone, J. F., Beecher, G., Holden, J., Haytowitz, D., et al. (2004).
Concentrations of proanthocyanidins in common foods and estimations of normal consump-
tion. Journal of Nutrition, 134 (3), 613–617.
10. Santos Buelga, C., & Scalbert, A. (2000). Proanthocyanidins and tannin like compounds –
Nature, occurrence, dietary intake and effects on nutrition and health. Journal of Agricultural
and Food Chemistry, 80 (7), 1094–1117.
References
68
11. Saura-Calixto, F., Serrano, J., & Goñi, I. (2007). Intake and bioaccessibility of total polyphe-
nols in a whole diet. Food Chemistry, 101 (2), 492–501.
12. Wang, Y., Chung, S. J., Song, W. O., & Chun, O. K. (2011). Estimation of daily proanthocyani-
din intake and major food sources in the US diet. Journal of Nutrition, 141 (3), 447–452.
13. Cassidy, A., Mukamal, K. J., Liu, L., Franz, M., Eliassen, A. H., & Rimm, E. B. (2013). High
anthocyanin intake is associated with a reduced risk of myocardial infarction in young and
middle-aged women. Circulation, 127 (2), 188–196.
14. Mazza, G. (2007). Anthocyanins and heart health. Annali dell’Istituto Superiore di Sanità,
43 (4), 369.
15. Passamonti, S., Vrhovsek, U., Vanzo, A., & Mattivi, F. (2003). The stomach as a site for antho-
cyanins absorption from food. FEBS Letters, 544 (1), 210–213.
16. Talavéra, S., Felgines, C., Texier, O., Besson, C., Lamaison, J. L., & Rémésy, C. (2003).
Anthocyanins are effi ciently absorbed from the stomach in anesthetized rats. Journal of
Nutrition, 133 (12), 4178–4182.
17. Passamonti, S., Vrhovsek, U., & Mattivi, F. (2002). The interaction of anthocyanins with bili-
translocase. Biochemical and Biophysical Research Communications, 296 (3), 631–636.
18. Talavéra, S., Felgines, C., Texier, O., Besson, C., Manach, C., Lamaison, J. L., et al. (2004).
Anthocyanins are effi ciently absorbed from the small intestine in rats. Journal of Nutrition,
134 (9), 2275–2279.
19. Prior, R. (2004). Absorption and metabolism of anthocyanins: Potential health effects. In M. S.
Meskin, W. R. Bidlack, A. J. Davies, D. S. Lewis, & R. K. Randolph (Eds.), Phytochemicals:
Mechanisms of action (pp. 1–19). Boca Raton: CRC.
20. He, J., Wallace, T. C., Keatley, K. E., Failla, M. L., & Giusti, M. M. (2009). Stability of black
raspberry anthocyanins in the digestive tract lumen and transport effi ciency into gastric and
small intestinal tissues in the rat. Journal of Agricultural and Food Chemistry, 57 (8),
3141–3148.
21. He, J., Magnuson, B. A., & Giusti, M. M. (2005). Analysis of anthocyanins in rat intestinal
contents impact of anthocyanin chemical structure on fecal excretion. Journal of Agricultural
and Food Chemistry, 53 (8), 2859–2866.
22. He, J., Magnuson, B. A., Lala, G., Tian, Q., Schwartz, S. J., & Giusti, M. M. (2006). Intact
anthocyanins and metabolites in rat urine and plasma after 3 months of anthocyanin supple-
mentation. Nutrition and Cancer, 54 (1), 3–12.
23. Felgines, C., Talavéra, S., Gonthier, M. P., Texier, O., Scalbert, A., Lamaison, J. L., et al.
(2003). Strawberry anthocyanins are recovered in urine as glucuro-and sulfoconjugates in
humans. Journal of Nutrition, 133 (5), 1296–1301.
24. Kroon, P. A., Clifford, M. N., Crozier, A., Day, A. J., Donovan, J. L., Manach, C., et al. (2004).
How should we assess the effects of exposure to dietary polyphenols in vitro? American
Journal of Clinical Nutrition, 80 (1), 15–21.
25. Gee, J. M., DuPont, M. S., Day, A. J., Plumb, G. W., Williamson, G., & Johnson, I. T. (2000).
Intestinal transport of quercetin glycosides in rats involves both deglycosylation and interac-
tion with the hexose transport pathway. Journal of Nutrition, 130 (11), 2765–2771.
26. Kay, C. D., Mazza, G. J., & Holub, B. J. (2005). Anthocyanins exist in the circulation primarily
as metabolites in adult men. Journal of Nutrition, 135 (11), 2582–2588.
27. Wallace, T. C. (2011). Anthocyanins in cardiovascular disease. Advances in Nutrition, 2 (1),
1–7.
28. Talavéra, S., Felgines, C., Texier, O., Besson, C., Gil-Izquierdo, A., Lamaison, J. L., et al.
(2005). Anthocyanin metabolism in rats and their distribution to digestive area, kidney, and
brain. Journal of Agricultural and Food Chemistry, 53 (10), 3902–3908.
29. Manach, C., Williamson, G., Morand, C., Scalbert, A., & Rémésy, C. (2005). Bioavailability
and bioeffi cacy of polyphenols in humans. I. Review of 97 bioavailability studies. American
Journal of Clinical Nutrition, 81 (1), 230S–242S.
30. Milbury, P. E., Vita, J. A., & Blumberg, J. B. (2010). Anthocyanins are bioavailable in humans
following an acute dose of cranberry juice. Journal of Nutrition, 140 (6), 1099–1104.
5 Anthocyanins Absorption and Metabolism
69
31. Pérez-Vicente, A., Gil-Izquierdo, A., & García-Viguera, C. (2002). In vitro gastrointestinal
digestion study of pomegranate juice phenolic compounds, anthocyanins, and vitamin C.
Journal of Agricultural and Food Chemistry, 50 (8), 2308–2312.
32. Ohnishi, R., Ito, H., Kasajima, N., Kaneda, M., Kariyama, R., Kumon, H., et al. (2006).
Urinary excretion of anthocyanins in humans after cranberry juice ingestion. Bioscience
Biotechnology and Biochemistry, 70 (7), 1681–1687.
33. Keppler, K., & Humpf, H.-U. (2005). Metabolism of anthocyanins and their phenolic degrada-
tion products by the intestinal microfl ora. Bioorganic and Medicinal Chemistry, 13 (17),
5195–5205.
34. Plumb, G. W., Price, K. R., & Williamson, G. (1999). Antioxidant properties of fl avonol gly-
cosides from green beans. Redox Report, 4 (3), 123–127.
35. Aura, A. M., Martin-Lopez, P., O’Leary, K. A., Williamson, G., Oksman-Caldentey, K. M.,
Poutanen, K., et al. (2005). In vitro metabolism of anthocyanins by human gut microfl ora.
European Journal of Nutrition, 44 (3), 133–142.
36. Schneider, H., & Blaut, M. (2000). Anaerobic degradation of fl avonoids by Eubacterium ram-
ulus. Archives of Microbiology, 173 (1), 71–75.
37. Fleschhut, J., Kratzer, F., Rechkemmer, G., & Kulling, S. E. (2006). Stability and biotransfor-
mation of various dietary anthocyanins in vitro. European Journal of Nutrition, 45 (1), 7–18.
38. Wu, X., Cao, G., & Prior, R. L. (2002). Absorption and metabolism of anthocyanins in elderly
women after consumption of elderberry or blueberry. Journal of Nutrition, 132 (7),
1865–1871.
39. Kalt, W., Blumberg, J. B., McDonald, J. E., Vinqvist-Tymchuk, M. R., Fillmore, S. A., Graf,
B. A., et al. (2008). Identifi cation of anthocyanins in the liver, eye, and brain of blueberry-fed
pigs. Journal of Agricultural and Food Chemistry, 56 (3), 705–712.
40. Andres-Lacueva, C., Shukitt-Hale, B., Galli, R. L., Jauregui, O., Lamuela-Raventos, R. M., &
Joseph, J. A. (2005). Anthocyanins in aged blueberry-fed rats are found centrally and may
enhance memory. Nutritional Neuroscience, 8 (2), 111–120.
41. McGhie, T. K., Ainge, G. D., Barnett, L. E., Cooney, J. M., & Jensen, D. J. (2003). Anthocyanin
glycosides from berry fruit are absorbed and excreted unmetabolized by both humans and rats.
Journal of Agricultural and Food Chemistry, 51 (16), 4539–4548.
42. Ichiyanagi, T., Shida, Y., Rahman, M. M., Hatano, Y., & Konishi, T. (2006). Bioavailability
and tissue distribution of anthocyanins in bilberry ( Vaccinium myrtillus L.) extract in rats.
Journal of Agricultural and Food Chemistry, 54 (18), 6578–6587.
43. Walle, T. (2004). Absorption and metabolism of fl avonoids. Free Radical Biology and
Medicine, 36 (7), 829–837.
44. Forester, S. C., & Waterhouse, A. L. (2008). Identifi cation of Cabernet Sauvignon anthocyanin
gut microfl ora metabolites. Journal of Agricultural and Food Chemistry, 56 (19), 9299–9304.
45. Forester, S. C., & Waterhouse, A. L. (2009). Metabolites are key to understanding health
effects of wine polyphenolics. Journal of Nutrition, 139 (9), 1824S–1831S.
46. Forester, S. C., & Waterhouse, A. L. (2010). Gut metabolites of anthocyanins, gallic acid,
3-O-methylgallic acid, and 2, 4, 6-trihydroxybenzaldehyde, inhibit cell proliferation of Caco-2
cells. Journal of Agricultural and Food Chemistry, 58 (9), 5320–5327.
47. Ávila, M., María, H., Concepción, S. M., Carmen, P., Teresa, R., & Pascual-Teresa, S. D.
(2009). Bioconversion of anthocyanin glycosides by Bifi dobacteria and Lactobacillus . Food
Research International, 42 (10), 1453–1461.
48. Azzini, E., Vitaglione, P., Intorre, F., Napolitano, A., Durazzo, A., Foddai, M. S., et al. (2010).
Bioavailability of strawberry antioxidants in human subjects. British Journal of Nutrition,
104 (08), 1165–1173.
49. Vitaglione, P., Donnarumma, G., Napolitano, A., Galvano, F., Gallo, A., Scalfi , L., et al.
(2007). Protocatechuic acid is the major human metabolite of cyanidin-glucosides. Journal of
Nutrition, 137
(9), 2043–2048.
References
71
© The Author(s) 2016
M. Riaz et al., Anthocyanins and Human Health, SpringerBriefs in Food,
Health, and Nutrition, DOI 10.1007/978-3-319-26456-1_6
Chapter 6
Biosynthesis and Stability of Anthocyanins
6.1 Introduction
Anthocyanins biosynthesis is one of the most studied secondary metabolite pathway
in plants. It is well-known now that anthocyanins biosynthesis is regulated by sev-
eral metabolic factors, majority of which are similar across various plant species.
Their biosynthetic pathway has been characterized at enzymatic as well as genetic
level with gene sequences available for all major biosynthetic steps of anthocyanins.
Further various mechanisms that regulate expression of these genes in plant cells
have also been studied.
Figure 6.1 summarizes the main steps of anthocyanins biosynthetic pathway. The
biosynthesis (phenylpropanoid metabolic pathway) starts from phenylalanine, an
amino acid, which yields 4-coumaroyl-CoA. This compound is then pooled with
malonyl-CoA to produce chalcones comprising two phenyl rings. The conjugate
ring-closure forms the chalcones, the well-known structure of fl avonoids, repre-
sented by the three-ringed structure (fl avanones). Flavanones are subjected to a
series of enzymatic modifi cations that produces dihydrofl avonols, anthocyanins but
also fl avonols, fl avan-3-ols, proanthocyanidins (tannins) and other polyphenolics.
The dihydro-fl avonols are converted to fl avonols (myricetin) which further are gly-
cosylated by glycosyl-transferase to the corresponding anthocyanins. Based on the
known biosynthetic pathways of anthocyanins, optimal intervention on the involved
enzymes of the secondary metabolic pathways may lead to an increase of anthocy-
anin content of the plant, as shown by research on strawberries [ 3 ].
Anthocyanins are produced from precursors by two biosynthetic pathways:
shikimate, producing phenylalanine; and that generating malonyl-CoA. These two
precursors are interconnected by chalcone synthase by a polyketide folding mecha-
nism, to yield an intermediate chalcone, a substrate for chalcone isomerase
producing prototype pigment naringenin which is oxidized by a series of enzymes
(fl avanone 3-hydroxylase, fl avonoid 3′-hydroxylase and fl avonoid 3′, 5′-hydroxylase)
to leucoanthocyanidins, and fi nally converted to anthocyanidins by
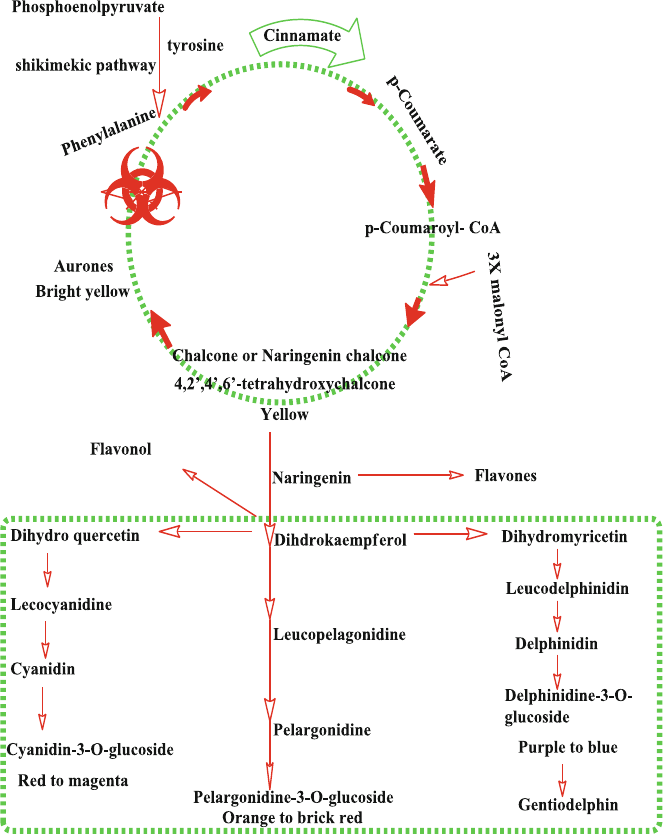
72
leucoanthocyanidin- dioxygenase. These unstable anthocyanidins are joined to
various sugars by UDP-glucose/fl avonoid 3- O -glucosyltransferase and O -methyl-
transferase to yield anthocyanins [ 4 ]. They accumulate in vacuoles of cells and
tissues of both reproductive and vegetative organs of plants. Out of 17 isolated
anthocyanins, majority are found in reproductive organs while 6 aglycones have
been identifi ed in vegetative parts of plants. Enzymes involved in biosynthetic
Fig. 6.1 Anthocyanins biosynthetic pathway [ 1 , 2 ]
6 Biosynthesis and Stability of Anthocyanins

73
pathway of anthocyanins are localized on the cytosolic side of smooth endoplasmic
reticulum (SER). From SER, they are transported to cell vacuole by multidrug and
toxic compound extrusion (MATE) and ATP-binding cassette (ABC) membrane
transported and vesicles.
6.2 Stability of Anthocyanins
Stability of anthocyanins is a major topic of recent research on anthocyanins due to
their potential applications, benefi cial effects and their use as alternative to artifi cial
colors. More detailed discussion is available about the stability of anthocyanins in
their concerned chapter however in product formulation this point may be consid-
ered that the stability of anthocyanins increases with increasing the viscosity of their
solution and molecular associations within the solution also increase the stability by
preventing water based degradations in anthocyanins. Anthocyanins are extremely
unstable and very prone to degradation. Their stability depends on oxygen, light,
temperature, pH, chemical structure, concentration, solvents, and presence of metal
ions, enzymes, proteins and fl avonoids [ 5 – 8 ]. Table 6.1 includes composition of
various anthocyanins.
Cyanidin and delphinidin are more stable than malvidin, peonidin and petuni-
din due to the blocking reactive OH group by methylation. Similarly anthocya-
nins with a 4-substituition are more stable than others. Glycosylation of 3
positions leads to stability while glycosylation of 5 position decreases stability.
Acylation of anthocyanins increases their stability by self-association reactions
and intra-molecular and intermolecular copigmentation. Therefore acylated
anthocyanins can offer the necessary stability for food applications. Acylated
anthocyanins can be obtained from black carrots, red cabbage, radish, purple
sweet potatoes and red potatoes [ 5 ].
Table 6.1 Edible sources of acylated anthocyanins [ 5 ]
Source Type of pigments
Radish ( Raphanus sativus ) Pelargonidin derivative acylated with one cinnamic acid and an
aliphatic acid
Potato ( Solanum tuberosum ) Pelargonidin derivatives acylated with one cinnamic acid
Black carrot ( Daucus carota
L.)
Cyanidin-3-Rutinoside-Glucoside-Galic acid acylated with one
cinnamic acid
Red cabbage ( Brassica
oleracea )
Cyanidin-3-Diglucoside-5-Glucoside acylated with one or two
cinnamic acids
Grape ( Vitis labrusca ) A mixture of fi ve different aglycones, acylated and non-acylated
with p-coumaric acid
6.2 Stability of Anthocyanins
74
6.3 Relationships Between Structure and Stability
Anthocyanins are often partially degraded under the joint action of cellular and
environmental factors (e.g. light, temperature, pH, metal ions, oxygen, coexisting
sugar, etc.). Their molecular structure (e.g., the number and placement of the
hydroxyl and methoxyl groups) affects their chemical behavior. The presence of
hydroxyl groups on the rings makes the anthocyanins carry a positive charge in
acidic solution. Although, the increased hydroxylation could stabilize the anthocy-
anidin, but excessive hydroxyl groups could also decrease the stability of the pig-
ment molecule, which makes it less stable than those with more methoxyl groups.
In the glycosides of anthocyanidin, the types of glycosyl units and acyl groups
attached to the aglycone have a signifi cant effect on the structural stability together
with their bonding site and number. Acylated anthocyanins are stable due to piling
of acyl groups with the pyrylium ring of the fl avylium cation, thus protecting the
chromophores from nucleophile attack of water leading to synthesis of chalcone or
pseudo-base [ 5 ]. This idea was also supported by Amiot and his coworkers [ 9 ], this
phenomenon is termed as intramolecular copigmentation. The color stability is
attributed in unacylated anthocyanins to intermolecular copigmentation mechanism
(including π–π overlap, dipole–dipole and hydrogen bonding interactions) [ 7 , 10 ].
Table 6.2 presents a comparative stability of various anthocyanins.
Table 6.2 Comparative stability of anthocyanins [ 10 ]
More Stable Stable Reference
Pyranoanthocyanins Anthocyanins > Anthocyanidins [
5 , 11 ]
Disaccharide anthocyanins Monosaccharidic anthocyanins [
5 , 11 ]
Acylatedanthocyanins unacylated derivatives
anthocyanins
[
5 , 11 ]
Polyacylatedanthocyanins Monoacylated anthocyanins [
5 , 11 ]
Anthocyaninsacylated with aromatic acids
(e.g. p-coumaric, caffeic, ferulic acids)
Acylated with aliphatic acids (e.g.
acetic, malonic or oxalic acids)
[
5 , 11 ]
Acylatedanthocyanins with caffeic acid Acylated anthocyanins with
p-coumaric acid
[
5 , 11 ]
aromatic acids substitution in ring B of
fl avyliumcation
aromatic acids substitution in ring
A of fl avylium cation
[
12 ]
Color of anthocyanins with petunidin or
malvidinaglycones
Color of anthocyanins with
pelargonidin, cyanidin, or
delphinidinaglycones
[
13 ]
Anthocyanins containing galactose Anthocyanins containing arabinose [
13 ]
Anthocyanins at low pH in acidic media Anthocyaninswith high pH in
alkaline solution
[
6 ]
6 Biosynthesis and Stability of Anthocyanins
75
6.4 Factor Affecting Stability of Anthocyanins
6.4.1 pH
Anthocyanins are susceptible toward change in pH [ 14 ] in media thus different
chemical forms and colors [ 15 ]. In acidic aqueous solutions, anthocyanins occur in
4 equilibrium species: the quinonoidal base (QB), the fl avylium cation (FC), the
carbinol or pseudo-base (PB) and the chalcone (CH) [ 16 , 17 ]. Table 6.3 indicates
the effect of pH on anthocyanins.
Anthocyanidins are less stable at neutral pH while Pg is the most stable anthocy-
anidin among all, in anthocyanins, monoglycosides and diglycosides derivatives are
more stable at pH 7 [ 18 ]. It is concluded that color variation with pH is more impor-
tant in the alkaline region due to their instability [ 7 ].
Ozela et al. (2007) evaluated the stability of anthocyanin in the extract of spinach
vine fruit ( Basella rubra L.) in relation to degradative factors such as light, tempera-
ture and pH acting alone or in combination. Spinach vine extract is more stable at
pH = 5.0–6.0 than at pH 4.0, in the absence as well as presence of light. This feature
is different from other anthocyanins. This property indicates its potential use as a
natural food color [ 19 ].
6.4.2 Co-pigmentation Effect
The anthocyanins color and intensity effects are the result of forming molecular or
complex associations is a co-pigmentation phenomenon for anthocyanins [ 20 ].
Some investigators propose it as stabilization mechanism for anthocyanins color in
plants [ 21 ]. The electron defi cient fl avylium ion associate with p-electrons rich co-
pigment thus gives stability against nucleophilic attack of water on 2 position of
fl avylium [ 22 ] and for other species such as SO
2
and peroxides on 4 position [ 23 ].
Figure 6.2 indicates anthocyanin interactions.
The co-pigmentation effect is pH dependent, because at low pH, anthocyanins
occur in fl avylium form, and at high pH values, they exist in colorless carbinol pseu-
dobase form. The example of charge transfer complex or p–p complex is the antho-
cyanins and tannins complexation in wines, producing pigmented tannins [
24 ].
Similar reactions occur between fl avan-3-ols and tannins [
7 , 25 ].
Metals and fl avylium salts chelates were considered for variety of colors in
fl owers [ 26 ]. Anthocyanin-metal complexations constitutes possible substitute for
color stabilization especially if the metals have no toxic effects. The complex for-
mation occurs with o-di-hydroxyl groups in the B ring (Cy, Dp, Pt) of anthocyanins
and anthocyanidins [ 20 ]. Some authors suggest that the blue color in plants is due
to complexation between anthocyanins and certain metals like Al, Fe, Cu and Sn
[ 27 ] or Mg and Mo [ 28 ]. Anthocyanin-molybdenum complexation causes the
6.4 Factor Affecting Stability of Anthocyanins
76
Table 6.3 Effect of pH on anthocyanins [ 7 ]
pH of the medium Predomanint chemical form Structure
1 Flavylium cation (red color)
O
HO
O
R
1
R
2
OH
O
gly
gly
Between 2 and 4 Quinoidal blue species
O
HO
O
R
1
R
2
O
O
gly
gly
Between 5 and 6 Carbinolpseudobase and a chalcone (colorless)
O
HO
O
R
1
R
2
OH
O
gly
gly
OH
HO
O
R
1
R
2
OH
O
gly
gly
O
OH
Chalcone
Psedobase carbinol
>7 Corresponding degraded products
OH
OH
Pyrocatechol
6 Biosynthesis and Stability of Anthocyanins

77
stabilization of blue color in Hindu cabbage tissue [ 28 ]. Yoshida et al. (2006) also
confi rmed the role of metal complex in plants by complexation between anthocya-
nins and proved that Mg (II) or Fe (III) at pH 5 are important for the development
of blue color in plants [ 7 , 29 ].
6.4.3 Solvent Effects
Studies on synthetic fl avylium salts in different solvents (acetonitrile, dioxane, etha-
nol, propylene-glycol, water and 2-butanone) have shown that variations in color
depend on the fl avylium salts concentration and solvent used [ 30 ]. In protic sol-
vents, the fl avylium salts display red color due to monomer, while in aprotic sol-
vents the solutions are yellow due to dimer while increasing concentration of
fl avylium salts, the red color is preferred. Another study suggests that increasing
H
2
O concentration in mixture of water:acetonitrile, the monomer is changed to a
green color dimer (monomer with charge-transfer property). Water therefore plays
an important role in dimerization of fl avylium salts [ 7 ].
ANs-
copigment-
interactions
Charge-transfer
complex
formation e.g.
another phenolic
compound
Self association
or an
intramolecular
co-pigmentation
e.g. other
anthocyanin
More complex
co-pigmintation
e.g. aglycon,
sugar, co-
pigment and
protons
Intermolecular
co-pigmentation
e.g. free
electrons
Compelexation
e.g., metals
Fig. 6.2 Anthocyanins (ANs) interaction [ 7 ]
6.4 Factor Affecting Stability of Anthocyanins
78
6.4.4 Temperature
The increase in temperature cause the decrease in stability of anthocyanins and
other plant pigments during processing and storage [ 8 ]. Rise in temperature cause
thermal degradation which yields brown products in the presence of oxygen [ 31 ].
Various researchers have confi rmed that heating has detrimental effect on anthocya-
nins stability [ 32 – 34 ].
6.4.5 Concentration Effects
Anthocyanin concentration has direct effect on color stability [ 5 ] such as strawberry
syrup color was greatly enhanced by mounting the anthocyanins contents. Total
anthocyanins are more color stabilizing than individual anthocyanins [ 35 ].
6.4.6 Oxygen
The presence of oxygen amplifi ed the degradation process, and the condition is
worsening if heating is there. Oxygen and increased temperature were the most
harmful combinations ever found in color degradation of various berry juices and
isolated anthocyanins [ 36 ] demonstrated that oxygen-free environment remained
best condition for the antioxidant activity and anthocyanins concentrations of straw-
berries (fresh-cut) during cold storage. Some investigators have reported that if the
atmosphere is enriched with 60–100 % oxygen results increase in phenolic and
anthocyanin content during the onset (0–7 days) of cold storage [ 37 ]. Yellow col-
ored 3-deoxyanthocyanins (due to dehydroxylation of the carbon at C-3) are more
stable than the corresponding red colored 3-hydroxy anthocyanins.
6.4.7 Light
Light is crucial for the biosynthesis of anthocyanins, but it also speeds up their deg-
radation. Dark has safeguarding effect on anthocyanins in comparison to light [ 38 ].
Amr and Al-Tamimi [
39 ] in 2007 studied the effect of light on anthocyanins stabil-
ity; their work verifi ed that light had a very detrimental effect on anthocyanins stabil-
ity during storage particularly in the presence of sugar. Light exposure cause
degradation of anthocyanins in incoherent manner which is source (plant) and antho-
cyanins dependent degradation [ 40 , 41 ]. Another study under neon light showed that
the phenolic and anthocyanin content was decreased during storage [ 42 ].
6 Biosynthesis and Stability of Anthocyanins
79
Ozela et al. (2007) [ 19 ] evaluated that various factors like pH, temperature and
light affect the anthocyanins stability of the spinach vine fruit ( Basella rubra L.)
extract. Results showed that, independent of pH, the extract exposed to light have
anthocyanin degradation kinetics with the average half-life is smaller compared to
an average half-life in dark storage. They found that in the presence of light,
increased temperature leads to increased anthocyanin degradation.
6.4.8 Enzymes
Glycosidases are the most common anthocyanins degrading enzymes that break
the glucosidic linkage and results unstable anthocyanidin [ 43 ]. Other anthocya-
nins degrading enzymes like peroxidases (phenol oxidases) and phenolases (poly-
phenol oxidases), are present in berries and fruits [ 44 ]. Inactivation of enzymes
increases anthocyanin stability [ 45 ]. Fang et al. (2007) [ 46 ] reported that bayberry
phenolase alone cannot react with directly with cyaniding-3-glucoside but
Mathew and Parpia [ 47 ] and Kader et al. (1999) [ 44 ], demonstrated that polyphe-
nol oxidase can react directly with anthocyanins although they acted as fairly poor
substrates.
6.4.9 Ascorbic Acid
Anthocyanins are degraded in the presence ascorbic acid by direct condensation
mechanism [ 48 ] and by free radical mechanism the pyrilium ring undergo oxidative
cleavage resulting in loss of color of anthocyanins. Ascorbic acid and its degrada-
tion products increase the degradation arte of anthocyanins [ 49 ].
6.4.10 Sugars
The results of adding sugar to anthocyanin stability depends mainly on the the num-
ber and nature of sugars attached to the fl avylium cation and number and nature of
acids linked to the glycosylic moiety [ 50 ]. Sugars and their degradation products
decrease anthocyanin stability. The addition of sucrose to mulberry extract increases
the stability of anthocyanins by increase in polymeric and co-pigmented anthocya-
nins during heating before heating the effect was hyperchromic [ 51 ]. Similarly
anthocyanins content of refrigerated berries extracts were more stabilized by 20 %
sugar addition however by increasing the concentration the protective action was
decreased [ 52 ].
6.4 Factor Affecting Stability of Anthocyanins
80
6.4.11 Sulfi tes
Sulphates and sulfi tes in foods items form colorless sulfur derivatives with antho-
cyanins which may be reversed by heating or acidifi cation if the concentration of
sulfi tes are not too high (>10 g/kg) [ 53 ].
6.5 Stability of Anthocyanins in Food Products
Due to plentiful applications, valuable effects and their use as alternative to artifi cial
colorants in foods; anthocyanins are the main focus of recent investigations. Food
processing involves extreme temperature up to 150 °C that may affect the stability
of anthocyanins during shelf life. Apart from temperature, other factors like pH,
storage temperature, chemical structure, concentration of anthocyanins present,
light, oxygen, the presence of enzymes, proteins and metallic ions also affect the
shelf lives of anthocyanins enriched products [ 54 ]. Generally high temperature may
affect concentration of anthocyanins in corresponding food items and products,
elevated temperature results in formation of polyphenolic degradation products that
may have low antioxidant potentials. Studies are required to confi rm whether these
thermal induced degradation products have decreased or increased antioxidant
capacity. Since the degradation mechanism of anthocyanins is rather complex and
perplexing, it is possible that thermal processing could induce some un/expected
and un/desired chemical reactions which in directly infl uence food quality so fur-
ther investigations are required in this area to predict optimum processing condi-
tions and to opt advance processing rather conventional system for respective
anthocyanins products to ensure maximum anthocyanins concentration, and longer
shelf lives products to the consumers [ 54 ].
6.6 Anthocyanins Degradation in Plants
Anthocyanins degradation is controlled and induced when benefi cial to the plant.
Anthocyanins degradation is dependent on environmental conditions such as light
quality, light intensity and growth temperature. Several enzymes have also been
isolated that degrade anthocyanins in postharvest fruit that may be candidates for
in vivo degradation. Peroxidase, α-glucosidase and polyphenol oxidase are the main
enzyme groups that regulate anthocyanins degradation in fruit juices and extracts
[ 55 ]. All research studies whether carried out in model systems or in extracts show
that anthocyanin degradation follows fi rst order kinetics.
Anthocyanins usually accumulate in young leaves and degrade as the leaves
become mature [ 56 ]. Anthocyanins protect the leaves especially their photosynthetic
6 Biosynthesis and Stability of Anthocyanins
81
apparatus from harmful UV light as well as from photo-inhibitory high intensities of
visible light [ 57 ]. They also act as antioxidants thus protecting the cells from oxida-
tive damage [ 58 ]. The decrease in anthocyanins content may be due to increase
chlorophyll accumulation as leaf expand and grow, and no longer is photoprotection
required [ 55 ].
Anthocyanins degradation is also observed in developing fruit e.g. Capsicum
spp. and Sicilian sweet orange varieties. Anthocyanins accumulation in fruit pro-
tects the photosynthetic apparatus in the developing fruit [ 55 ].
Flowers often change color during development, acting as a signal for pollina-
tors. In most cases, the change in color is due to induction of anthocyanins synthe-
sis, but in others, such as Brunfelsia calycina , anthocyanin is degraded, resulting in
a change of fl ower color from dark purple to white after synthesis [ 59 ].
6.7 Drawback and Derivatives of Anthocyanins
As previously discussed, anthocyanins are unstable and their colors varies depend-
ing upon light, temperature, pH and presence of metals in media [ 60 ]. There are
some limitations like decreased stability to processing, storage and formulation
conditions which can impart un-wanted fl avor or odor characteristics that have
limited their use as natural colorants in food systems. The area required attention
to developed processes and mechanisms to reduce the incompatibility of anthocya-
nins with pH, temperature and other environmental changes. For example there
may be few coloring principles in red cabbage and radish extracts so techniques
and procedures must be adopted to isolated these few necessary components as
coloring agents [ 5 ].
6.8 Anthocyanins Stabilization Mechanisms
As mentioned before, naturally anthocyanins are unstable, however they can be
stabilized i.e. by applying various mechanisms like acetylations, association (co-
pigmentation) and microencapsulation when a “co-pigment” is added. The stabili-
zation of grape skin anthocyanins by co-pigmentation with enzymatically modifi ed
isoquercitrin (EMIQ) was increased signifi cantly [ 61 ].
Encapsulation by spray-drying is a cost-effective way for protection of anthocya-
nins by coating the ingredients [ 62 ]. Thus bioavalability and the shelf lives of antho-
cyanins may be improved by providing protection against oxygen, water and light
[ 53 ]. The encapsulation from diverse sources is given in Table 6.4 and the details of
steps during process and their advantages are also given in Table 6.5 .
6.8 Anthocyanins Stabilization Mechanisms
82
Table 6.4 Anthocyanins encapsulation from various sources [
53 ]
Anthocyanin
source
Encapsulation
technique Coating material Stabilization improvement
Concord grape Spray drying Maltodextrin Not measured
Cranberry
pomace
Spray drying Maltodextrin Not measured
Roselle calyces Spray drying Maltodextrin Not measured
Roselle calyces Inclusion
complexation
β-cyclodextrin The addition of β-cyclodextrin
does not ↑ anthocyanin
stabilization
Apple pomace Spray drying Maltodextrin Anthocyanins have↑ stability
The ↑ in the shelf life is due to
↓ water activity
Roselle calyces Liophilization Pullulan Encapsulated anthocyanins have
slightly ↑ stability as compared
to the free anthocyanin extract
Black currant Thermal gelation Glucan gel Encapsulation of anthocyanins
followed by appropriate
processing ↑ the stability of
anthocyanins
Black carrot Spray drying Maltodextrin Not measured
‘Isabel’ grape
bagasse
Spray drying Maltodextrin and
Arabic gum
Not measured
Blackberry Thermal gelation Sodium alginate
and pectin
Not measured
Blackberry Ionic gelation Curdlan Not measured
Jabuticaba fruit
skin
Ionic gelation Sodium alginate Not measured
Table 6.5 Some common microencapsulation processes for anthocyanins characteristics [
63 ]
Technology
Particle
size
(μm) Advantages Disadvantages
Spray-drying 10–400 Low process cost, wide
choice of wall material,
good encapsulation
effi ciency, good stability of
the fi nished product and
potential of industrial
production continuously
Can degrade at highly
temperature-sensitive
compounds, control of the
particle size is diffi cult and
yields for small batches are
moderate
Freeze drying 20–
5000
Best for thermo-sensitive
substances in aqueous
solutions
More processing time, costly
process, costly transport and
storage of the capsules
Fluid bed coating 20–200 Low cost process and
allows specifi c capsule size
distribution and low
porosities into the product
Degradation of temperature-
sensitive molecules
(continued)
6 Biosynthesis and Stability of Anthocyanins
83
6.9 Conclusions
A lot of data is available on molecular biology of anthocyanin biosynthesis. The
genes regulating the various steps and the infl uence of phytohormones on this
genetic regulation are further clarifi ed now. Their intra-cellular transportation and
accumulation are also better understood. Although substantial progress has been
made in our understanding of anthocyanin biosynthesis in plants, many key issues
are yet to be addressed in detail. For example the interaction between anthocyanin
biosynthetic pathway and other pathways and the effects of such interaction on
plant growth and development are not fully understood. It is hoped that next decade
will clarify remaining ambiguities in this regard.
References
1. De Pascual-Teresa, S., & Sanchez-Ballesta, M. T. (2008). Anthocyanins: From plant to health.
Phytochemistry Reviews, 7 (2), 281–299.
2. Teixeira da Silva, J. A., Serena, A., Wei, L., Hao, Y., & Akira, K. (2014). Genetic control of
fl ower development, color and senescence of Dendrobium orchids. Scientia Horticulturae,
175 , 74–86.
Technology
Particle
size
(μm) Advantages Disadvantages
Emulsifi cation 0.2–
5000
Polar, non-polar (apolar),
and amphiphilic can be
incorporated
Low number of emulsifi ers that
can be utilized and hard to
control formation of capsules
Melt extrusion 300–
5000
The material is completely
shielded by the wall
material, remaining core is
washed externally, low
temperature entrapping
assay
The capsule should be
essentially separated from liquid
bath and dried
Hard to obtain capsules in highly
viscous carrier materials melts
Rapid expansion
of supercritical
fl uid (RESS)
10–400 Easy removal of solvent
and non-toxic method,
works at ↓temperature and
in in-active atmosphere
thereby avoiding product
degradation
Both the core and the wall
material must be very soluble in
supercritical fl uids, low or no
solubility of high molecular
weight, polar compounds CO
2
,
poor control over the
precipitated crystal morphology
and size distribution
Ionic Gelation Organic solvents and
extreme conditions of
temperature and pH are
avoided
Mostly used on lab-scale
The capsules are highly porous
which leads to intensive burst
Thermal Gelation The same of ionic gelation Same as mentioned above
Table 6.5 (continued)
References
84
3. Oancea, S., & Oprean, L. (2011). Anthocyanin extracts in the perspective of health benefi ts
and food applications. Revista de Economia, 218 .
4. Kassim, A., Poette, J., Paterson, A., Zait, D., McCallum, S., Woodhead, M., et al. (2009).
Environmental and seasonal infl uences on red raspberry anthocyanin antioxidant contents and
identifi cation of quantitative traits loci (QTL). Molecular Nutrition and Food Research, 53 (5),
625–634.
5. Giusti, M. M., & Wrolstad, R. E. (2003). Acylated anthocyanins from edible sources and their
applications in food systems. Biochemical Engineering Journal, 14 (3), 217–225.
6. Rein, M. (2005). Copigmentation reactions and color stability of berry anthocyanins . Helsinki:
University of Helsinki.
7. Castañeda-Ovando, A., Lourdes Pacheco-Hernández, M. D. L., Páez-Hernández, M. E., José,
A. R., & Galán-Vidal, C. A. (2009). Chemical studies of anthocyanins: A review. Food
Chemistry, 113 (4), 859–871.
8. Mercadante, A. Z., & Bobbio, F. O. (2008). Anthocyanins in foods: Occurrence and physico-
chemical properties. In C. Socaciu (Ed.), Food colorants: Chemical and functional properties
(pp. 241–276). Boca Raton: CRC.
9. Malien-Aubert, C., Dangles, O., & Amiot, M. J. (2001). Color stability of commercial antho-
cyanin-based extracts in relation to the phenolic composition. Protective effects by intra-and
intermolecular copigmentation. Journal of Agricultural and Food Chemistry, 49 (1), 170–176.
10. Li, H., Zeyuan, D., Honghui, Z., Chanli, H., Ronghua, L., Christopher, Y. J., et al. (2012).
Highly pigmented vegetables: Anthocyanin compositions and their role in antioxidant activi-
ties. Food Research International, 46 (1), 250–259.
11. Stintzing, F. C., & Carle, R. (2004). Functional properties of anthocyanins and betalains in
plants, food, and in human nutrition. Trends in Food Science and Technology, 15 (1), 19–38.
12. Yoshida, K., Reiko, O., Kiyoshi, K., & Tadao, K. (2002). Prevention of UV-light induced E,
Z-isomerization of caffeoyl residues in the diacylated anthocyanin, gentiodelphin, by intramo-
lecular stacking. Tetrahedron Letters, 43 (35), 6181–6184.
13. Von Elbe, J., & Schwartz, S. (1996). Colorants. Food Chemistry, 3 , 651–723.
14. Wesche-Ebeling, P., & Argaiz-Jamet, A. (2002). Stabilization mechanisms for anthocyanin the
case for copolymerization reactions. In J. Welti-Chanes, G. V. Barbosa-Cánovas, & J. M.
Aguilera (Eds.), Engineering and food for the 21st century (pp. 141–150). Boca Raton: CRC.
15. Brouillard, R., & Markakis, P. (1982). Anthocyanins as food colors . New York: Academic.
16. da Costa, C. T., Bryant, C. N., Sam, A. M., & Derek, H. (1998). Separation of blackcurrant
anthocyanins by capillary zone electrophoresis. Journal of Chromatography A, 799 (1),
321–327.
17. Kennedy, J. A., & Waterhouse, A. L. (2000). Analysis of pigmented high-molecular-mass
grape phenolics using ion-pair, normal-phase high-performance liquid chromatography.
Journal of Chromatography A, 866 (1), 25–34.
18. Fleschhut, J., Kratzer, F., Rechkemmer, G., & Kulling, S. E. (2006). Stability and biotransfor-
mation of various dietary anthocyanins in vitro. European Journal of Nutrition, 45 (1), 7–18.
19. Ferreira Ozela, E., Stringheta, P. C., & Cano Chauca, M. (2007). Stability of anthocyanin in
spinach vine ( Basella rubra ) fruits. Ciencia e Investigación Agraria, 34 (2), 115–120.
20. Boulton, R. (2001). The copigmentation of anthocyanins and its role in the color of red wine:
A critical review. American Journal of Enology and Viticulture, 52 (2), 67–87.
21. Davies, A., & Mazza, G. (1993). Copigmentation of simple and acylated anthocyanins with
colorless phenolic compounds. Journal of Agricultural and Food Chemistry, 41 (5), 716–720.
22. Matsufuji, H., Otsuki, T., Takeda, T., Chino, M., & Takeda, M. (2003). Identifi cation of reac-
tion products of acylated anthocyanins from red radish with peroxyl radicals. Journal of
Agricultural and Food Chemistry, 51 (10), 3157–3161.
23. Garcia-Viguera, C., & Bridle, P. (1999). Infl uence of structure on color stability of anthocya-
nins and fl avylium salts with ascorbic acid. Food Chemistry, 64 (1), 21–26.
24. Mirabel, M., Saucier, C., Guerra, C., & Glories, Y. (1999). Copigmentation in model wine
solutions: Occurrence and relation to wine aging. American Journal of Enology and Viticulture,
50 (2), 211–218.
6 Biosynthesis and Stability of Anthocyanins
85
25. Salas, E., Atanasova, V., Poncet-Legrand, C., Meudec, E., Mazauric, J. P., & Cheynier, V.
(2004). Demonstration of the occurrence of fl avanol-anthocyanin adducts in wine and in model
solutions. Analytica Chimica Acta, 513 (1), 325–332.
26. Clifford, M. N. (2000). Anthocyanins-nature, occurrence and dietary burden. Journal of the
Science of Food and Agriculture, 80 (7), 1063–1072.
27. Starr, M., & Francis, F. (1973). Effect of metallic ions on color and pigment content of cran-
berry juice cocktail. Journal of Food Science, 38 (6), 1043–1046.
28. Hale, K. L., McGrath, S. P., Lombi, E., Stack, S. M., Terry, N., Pickering, I. J., et al. (2001).
Molybdenum sequestration in brassica species. A role for anthocyanins? Plant Physiology,
126 (4), 1391–1402.
29. Yoshida, K., Kitahara, S., Ito, D., & Kondo, T. (2006). Ferric ions involved in the fl ower color
development of the Himalayan blue poppy, Meconopsis grandis. Phytochemistry, 67 (10),
992–998.
30. Ito, F., Nobuaki, T., Akio, K., & Tsuneo, F. (2002). Why do fl avylium salts show so various
colors in solution?: Effect of concentration and water on the fl avylium’s color changes. Journal
of Photochemistry and Photobiology A: Chemistry, 150 (1), 153–157.
31. Markakis, P. (2012). Anthocyanins as food colors . New York: Elsevier.
32. Jiménez, N., Bohuon, P., Lima, J., Dornier, M., Vaillant, F., & Pérez, A. M. (2010). Kinetics of
anthocyanin degradation and browning in reconstituted blackberry juice treated at high tem-
peratures (100−180°C). Journal of Agricultural and Food Chemistry, 58 (4), 2314–2322.
33. Lin, Y.-C., & Chou, C.-C. (2009). Effect of heat treatment on total phenolic and anthocyanin
contents as well as antioxidant activity of the extract from Aspergillus awamori -fermented
black soybeans, a healthy food ingredient. International Journal of Food Science and Nutrition,
60 (7), 627–636.
34. Sadilova, E., Florian, C., Stintzing, D. R., & Kammerer, R. C. (2009). Matrix dependent
impact of sugar and ascorbic acid addition on color and anthocyanin stability of black carrot,
elderberry and strawberry single strength and from concentrate juices upon thermal treatment.
Food Research International, 42 (8), 1023–1033.
35. Skrede, R. E. G., Wrolstad, P. L., & Enersen, G. (1992). Color stability of strawberry and
blackcurrant syrups. Journal of Food Science, 57 (1), 172–177.
36. Odriozola-Serrano, I., Soliva-Fortuny, R., & Martín-Belloso, O. (2010). Changes in bioactive
composition of fresh-cut strawberries stored under superatmospheric oxygen, low-oxygen or
passive atmospheres. Journal of Food Composition and Analysis, 23 (1), 37–43.
37. Zheng, Y., Shiow, Y. W., Chien, Y. W., & Wei, Z. (2007). Changes in strawberry phenolics,
anthocyanins, and antioxidant capacity in response to high oxygen treatments. LWT – Food
Science and Technology, 40 (1), 49–57.
38. Kearsley, M., & Rodriguez, N. (1981). The stability and use of natural colors in foods:
Anthocyanin, β-carotene and ribofl avin. International Journal of Food Science and Technology,
16 (4), 421–431.
39. Amr, A., & Al Tamimi, E. (2007). Stability of the crude extracts of Ranunculus asiaticus
anthocyanins and their use as food colorants. International Journal of Food Science and
Technology, 42 (8), 985–991.
40. Inami, O., Itaru, T., Hiroe, K., & Nobuji, N. (1996). Stability of anthocyanins of Sambucus
canadensis and Sambucus nigra. Journal of Agricultural and Food Chemistry, 44 (10),
3090–3096.
41. Shi, Z., Bassa, I. A., Gabriel, S. L., & Francis, F. J. (1992). Anthocyanin pigments of sweet
potatoes– Ipomoea batatas . Journal of Food Science, 57 (3), 755–757.
42. Maier, T., Matthias, F., Andreas, S., Dietmar, R., & Kammerer, R. C. (2009). Process and stor-
age stability of anthocyanins and non-anthocyanin phenolics in pectin and gelatin gels enriched
with grape pomace extracts. European Food Research and Technology, 229 (6), 949–960.
43. Huang, H. (1956). The kinetics of the decolorization of anthocyanins by fungal “anthocya-
nase” 1. Journal of the American Chemical Society, 78 (11), 2390–2393.
44. Kader, F., Irmouli, M., Zitouni, N., Nicolas, J. P., & Metche, M. (1999). Degradation of cyani-
din 3-glucoside by caffeic acid o-quinone. Determination of the stoichiometry and character-
ization of the degradation products. Journal of Agricultural and Food Chemistry, 47 (11),
4625–4630.
References
86
45. Garcia-Palazon, A., Suthanthangjai, W., Kajda, P., & Ioannis, Z. (2004). The effects of high
hydrostatic pressure on β-glucosidase, peroxidase and polyphenoloxidase in red raspberry
( Rubus idaeus ) and strawberry ( Fragaria ananassa ). Food Chemistry, 88 (1), 7–10.
46. Fang, Z., Min, Z., Yunfei, S., & Jingcai, S. (2007). Polyphenol oxidase from bayberry ( Myrica
rubra Sieb. et Zucc.) and its role in anthocyanin degradation. Food Chemistry, 103 (2),
268–273.
47. Mathew, A., & Parpia, H. (1971). Food browning as a polyphenol reaction. Advances in Food
Research, 19 , 75–145.
48. Poei‐Langston, M., & Wrolstad, R. (1981). Color degradation in an ascorbic acid anthocyanin
fl avanol model system. Journal of Food Science, 46 (4), 1218–1236.
49. Pacheco-palencia, L. A., Hawken, P., & Talcott, S. T. (2007). Juice matrix composition and
ascorbic acid fortifi cation effects on the phytochemical, antioxidant and pigment stability of
açai ( Euterpe oleracea Mart.). Food Chemistry, 105 (1), 28–35.
50. Rubinskiene, M., Viskelis, P., Jasutiene, I., Viskeliene, R., & Bobinas, C. (2005). Impact of
various factors on the composition and stability of black currant anthocyanins. Food Research
International, 38 (8), 867–871.
51. Tsai, P. J., Delva, L., Yu, T. Y., Huang, Y. T., & Dufosse, L. (2005). Effect of sucrose on the
anthocyanin and antioxidant capacity of mulberry extract during high temperature heating.
Food Research International, 38 (8), 1059–1065.
52. Nikkhah, E., Khayamy, M., Heidari, R., & Jamee, R. (2007). Effect of sugar treatment on
stability of anthocyanin pigments in berries. Journal of Biological Sciences, 7 (8),
1412–1417.
53. Cavalcanti, R. N., Santos, D. T., & Meireles, M. A. A. (2011). Non-thermal stabilization mech-
anisms of anthocyanins in model and food systems – An overview. Food Research International,
44 (2), 499–509.
54. Patras, A., Nigel, P. B., O’Donnell, C., & Tiwari, B. K. (2010). Effect of thermal processing on
anthocyanin stability in foods; mechanisms and kinetics of degradation. Trends in Food
Science and Technology, 21 (1), 3–11.
55. Oren-Shamir, M. (2009). Does anthocyanin degradation play a signifi cant role in determining
pigment concentration in plants? Plant Science, 177 (4), 310–316.
56. Chalker Scott, L. (1999). Environmental signifi cance of anthocyanins in plant stress responses.
Photochemistry and Photobiology, 70 (1), 1–9.
57. Steyn, W. J., Wand, S. J. E., Holcroft, D. M., & Jacobs, G. (2002). Anthocyanins in vegetative
tissues: A proposed unifi ed function in photoprotection. New Phytologist, 155 (3), 349–361.
58. Winkel-Shirley, B. (2002). Biosynthesis of fl avonoids and effects of stress. Current Opinion in
Plant Biology, 5 (3), 218–223.
59. Vaknin, H., Bar-Akiva, A., Ovadia, R., Nissim-Levi, A., Forer, I., Weiss, D., et al. (2005).
Active anthocyanin degradation in Brunfelsia calycina (yesterday–today–tomorrow) fl owers.
Planta, 222 (1), 19–26.
60. Da Costa, C. T., Horton, D., & Margolis, S. A. (2000). Analysis of anthocyanins in foods by
liquid chromatography, liquid chromatography-mass spectrometry and capillary electrophore-
sis. Journal of Chromatography A, 881 (1), 403–410.
61. Valentová, K., Vrba, J., Bancířová, M., Ulrichová, J., & Křen, V. (2014). Isoquercitrin:
Pharmacology, toxicology, and metabolism. Food and Chemical Toxicology, 68 , 267–282.
62. Cai, Y., & Corke, H. (2000). Production and properties of spray dried amaranthus betacyanin
pigments. Journal of Food Science, 65 (7), 1248–1252.
63. Mahdavi, S. A., Seid, M. J., Mohammad, G., & Elham, A. (2014). Spray-drying microencap-
sulation of anthocyanins by natural biopolymers: A review.
Drying Technology, 32 (5),
509–518.
6 Biosynthesis and Stability of Anthocyanins
87
© The Author(s) 2016
M. Riaz et al., Anthocyanins and Human Health, SpringerBriefs in Food,
Health, and Nutrition, DOI 10.1007/978-3-319-26456-1_7
Chapter 7
The Role of Anthocyanins in Health
as Antioxidant, in Bone Health and as Heart
Protecting Agents
7.1 Introduction
Anthocyanins are one of the most well-known sub-groups of pigmented fl avonoid
compounds in the plant kingdom. They can be found in almost all land plants, but
found in relatively high levels in 27 plant families. The dietary consumption of
anthocyanins is high due to their occurrence in fruits and vegetables. They function
as phytoprotective substances, have a role in plant, animal interactions and as such
are important in eco-physiology or plant defense mechanisms. Recently, interest in
anthocyanins as well as in their biological and therapeutic properties has strongly
increased. Nemours in vitro studies, animal models and human clinical trials sup-
port their medicinal benefi ts to human. These studies suggest that anthocyaninshave
anti-carcinogenic and anti-infl ammatory effects, provide cardiovascular disease
prevention, promote obesity and diabetes control benefi ts, and also improve visual
and brain functions. To understand the role of anthocyanins in prevention of dis-
eases, it is essential to have a working-knowledge of anti-oxidants and their regula-
tory role on the human immune system.
Reactive species is a collective term used to indicate oxygen radicals, nitrogen
radicals and their associated nonradicals species that act as oxidizing agents. These
species are known as reactive oxygen species (ROS) and reactive nitrogen species
(RNS) respectively or commonly as oxidants. These radicals can be produced by a
variety of mechanism in human body and their uncontrolled production can be
harmful for biomolecules, cells, tissues and body organs known as oxidative dam-
age. The in vivo generation of ROS and RNS elicits the radical scavenging or
antioxidant mechanism collectively known as antioxidant defense which minimizes
and repairs this oxidative damage. This oxidant-antioxidant balance is required for
proper functioning of cells and during disease; this balance is sloped in favor of
oxidants thus generating oxidative stress and increasing oxidative damage. The
ROS and RNS are involved in pathology of more than 100 various diseases and
metabolic disorders. This damage may be countered by oral administration of
88
antioxidants from antioxidant rich sources of fruits, vegetables, nuts, grains and
cereals. It has led to new antioxidant based preventive/curative nutrition based ther-
apies. These antioxidants modulate innate as well as adaptive immunity and reverse
various age-related immune defi ciencies by stimulating and boosting the human
immune system. Although more signifi cant effects have been noted in elderly, the
antioxidant rich diets have same effects in youngers. It is therefore recommended to
have a suffi cient intake of antioxidant rich foods regularly to prevent and delay the
onset of various age-related degenerative disorders. Some of these food-related
antioxidants are ascorbic acid, tocopherols, carotenoids and anthocyanins.
It is well-established now that anthocyanin stimulates immunity against tumor
growth. Anthocyanin intervention can be helpful strategy in modulating the immune
response. Whether the immune modulatory effects of anthocyanins translate into
health benefi ts should be studied with various biological and animal models. It must
be kept in mind that these studies have their drawbacks as well. For example, studies
using one pure anthocyanin, the aspect of interaction of various dietary bioactive
constituents and complete array of various anthocyanin are missing. Similarly for
studies involving plant material or extract, which contain collection of other
constituents and so results can’t be attributed to anthocyanin only.
Different signaling pathways like nuclear factor κB, Wnt/β-catenin, AMP-
activated and mitogen-activated protein kinase as well as certain central cellular
processes, such as cell cycle, apoptosis, autophagy and biochemical metabolism are
involved in these valuable effects of anthocyanins and can provide potential thera-
peutic strategies for treatment of broad spectrum of diseases in future [ 1 ]. Figure 7.1
gives a general overview of pharmacological effects of anthocyanins on various
organs and tissues of human body.
7.2 Presumed Health-Promoting Effects of Anthocyanins
Due to their health-promoting and immunity-boosting properties, anthocyanins are
appropriate nutraceuticals and supplementary treatments for various aspects of
chronic diseases. Dieto-therapeutic applications of anthocyanin-rich foods optimize
health and performance constituting an important largest market of nutraceutical
products throughout the world. Therefore, it is strongly recommended to identify
these health-promoting components in anthocyanin extracts that led to new opening
of opportunities of using these extracts in a variety of food applications.
A lot of research has been performed during last decade to identify non-essential
nutrients including anthocyanin responsible for observed health-benefi ts and to
verify various claimed physiological effects on human body. Their benefi cial effects
include anti-infl ammatory and anti-cancer activities and protective effects against
various metabolic, degenerative and cardiovascular diseases and vision improve-
ments. There are numerous studies in animal/human cell lines, animal models and
7 The Role of Anthocyanins in Health as Antioxidant, in Bone Health and as Heart…
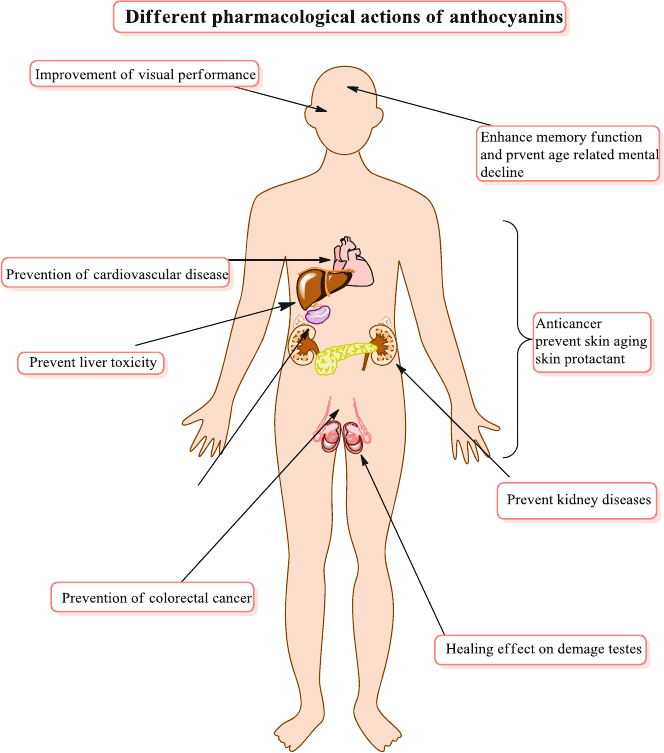
89
human clinical trials that support above mentioned effects of anthocyanins. Their
peculiar chemical diversity, suitable molecular weight, specifi c three dimensional
conformation and resulting physical and biochemical effects allow them to interact
with various targets in cells producing desirable effects. Data retrieved from various
studies support the health-promoting and immunity boosting effects of anthocya-
nins. Their and their metabolites role in maintaining optimal functions of various
physiological and biochemical processes is supported by epidemiological studies,
in vitro studies, cell-culture studies and recently in vivo studies. These efforts have
led to better understanding of the role of anthocyanin and their derivatives in the
Fig. 7.1 General Overview of pharmacological effects of anthocyanins
7.2 Presumed Health-Promoting Effects of Anthocyanins

90
process of various chronic diseases, with special focus to their metabolism and
biological actions, dose-effects and organ specifi c effects. Table 7.1 indicates
healthy effects of uses of anthocyanins on various tissues, organs and organ systems
of human body.
Interest in anthocyanins has increased after the recognition of their health bene-
fi ts [ 16 ]. Due to their anti-infl ammatory and anti-oxidant potential, epidemiologic
studies suggest that ingestion of anthocyanins reduces the risk of diabetes, arthritis,
cancer and cardiovascular disease [ 17 ]. Their effects are due to their effects at
Table 7.1 Healthy effects of uses of anthocyanins
Anthocyanins Health use References
Anthocyanins extracts Sight acuteness [
2 ]
Enhancement of antioxidant capacity [
3 , 4 ]
Cured several blood circulation ailments
due to capillary fragility
[
5 ]
Vaso-protective and anti-infl ammatory
effects
[
6 ]
Platelet aggregation inhibition effects [
7 ]
Preservation of normal vascular
permeability
[
5 ]
Anti-diabetic, anti-neoplastic and chemo-
protective effects
[
8 , 9 ]
Radiation-protective agents [
10 ]
Antioxidant properties and notable effects
against Cardiovascular hypertension,
chronic infl ammation, Cancer and metabolic
syndrome
[
11 , 12 ]
Red orange extract ( Citrus sinensis
varieties: Moro, Tarocco,
Sanguinello) (Anthocyanins,
fl avanones, hydroxycinnamic
acids, and ascorbic acid)
Anti-infl ammatory activity was assessed in
human keratinocytes (lineage NCTC 2544)
exposed to IFN-γ and histamine
[
13 , 14 ]
Red orange extract at various
concentrations
Halted expression of ICAM-1 and secretion
of MCP-1 and IL-8
[
13 , 14 ]
Grape seed proanthocyanidins Human keratinocytes irradiated with UVB
and treated with GSP’s inhibited formation
of UVB-induced H
2
O
2
, protein oxidation,
lipid peroxidation, DNA damage and
exhaustion of antioxidant components like
catalase, superoxide dismutase, glutathione
peroxidase and glutathione
[
14 , 15 ]
Grape seed proanthocyanidins Prevented phosphorylation of JNK, p38,
ERK1/2and proteins of MAPK family and
UVB-induced activation of NF-κB/p65. It
suggests that GSP canreduce UV-induced
oxidative stress in human skin
[
14 , 15 ]
7 The Role of Anthocyanins in Health as Antioxidant, in Bone Health and as Heart…

91
various molecular levels by directly scavenging ROS [ 18 ] or inducing phase II
detoxifying enzymes or to induce phase II detoxifying enzymes [ 19 ].
7.3 Toxicity of Anthocyanins
No adverse health effects have been associated with anthocyanins consumption
[ 20 , 21 ]. Even if consumption in large quantities in countries of Europe and USA,
no toxicity has been observed, probably due to their low overall absorption.
However the over-increasing usage as supplements may prove problematic. In tox-
icity studies in animal models, using bilberry extracts (36 % anthocyanindins),
LD
50
values were over 2000 mg/kg without any toxic indications [ 22 ]. Similarly in
dogs, no mortality or toxic effects were observed at 3000 mg/kg. No adverse or
toxic effects were observed in rats (@ 125–500 mg/kg) and dogs (@80–320 mg/kg)
daily for 6 months. The extract did not exhibit any teratogenic or mutagenic effects
thus confi rming clinical safety studies [ 22 ]. People consuming 160 mg/kg twice
daily for 1–2 month tolerated the extract very-well. Only some (4 %) complained
side effects, mostly gastrointestinal (GIT) and nervous system and cutaneous
annexes and skin [ 23 ].
Anthocyanins have been consumed since ancient times but no toxic effects
regarding health were reported [ 24 ]. That is the reason that Europe, Japan, USA
and many other countries allowed the use of anthocyanins as food color in
foods and beverages [ 25 ]. The Joint FAO/WHO Expert Committee on Food
Additives (JECFA) determined on early toxicological studies that anthocyanin-
containing extracts had a very low toxicity [ 26 ]. The no-observed-effect-level
(NOEL) for young rats was determined to be approximately 225 mg kg
−1
body
weight in a two-generation reproduction study. Based on the above result, the
estimated acceptable daily intake (ADI) for human was estimated to be
2.5 mg kg
−1
body weight per day in 1982, using the equation of ADI = NOEL/100
[ 27 ]. The studies presented in Tables 7.1 and 7.2 have confirmed the safety of
anthocyanins.
Table 7.2 Various mutagenicity studies are described by JECFA [ 28 ]
Anthocyanins Model tested Genotoxicity References
Cyanidin and
delphinidin
Ames assay system for fi ve different
strains of Salmonella typhimurium
Not mutagenic [
29 ]
Cyanidin Ames assay using Salmonella
typhimurium strain TA98
Not mutagenic [
30 ]
Anthocyanins
(compounds not
specifi ed)
Ames test using Salmonella
typhimurium TA1538
Not mutagenic [
31 ]
Anthocyanins
(compounds not
specifi ed)
Escherichia coli WP2 for induction of
DNA damage
Not mutagenic [
31 ]
7.3 Toxicity of Anthocyanins
92
7.4 Reproductive and Developmental Toxicity
Grape-skin extract containing 3 % anthocyanins were evaluated for 2-generation
reproduction study in Sprague–Dawley rats confi rmed its safety in reproduction and
growth; there were no corresponding compound-related histopathological effects
[ 32 ]. Anthocyanins (an extract from currants, blueberries and elderberries) were
reported not to be a developmental toxicant in rats, mice or rabbits when given over
three successive generations [ 28 ]. Table 7.3 shows effects of processing conditions
on anthocyanin contents (mg/g DW) of fruits.
7.5 Anthocyanins Biological and Pharmacological Activities
Anthocyanins are not only benefi cial to its producers, i.e. plants but also to humans and
other animals as nutraceutical and coloraceutical. In present era numerous scientists
have focused anthocyanins, most of them have found anthocyanins as useful and
benefi cial reporting wide range of biological activities [ 11 ]. These activities include
anti-infl ammatory [ 34 ] antimicrobial [ 35 ] anti-carcinogenic [ 11 , 36 ] vision- improvement
[ 37 ], stimulation of apoptosis [ 36 ]; neuroprotective effects [ 38 , 39 ] effects on blood
vessels [ 40 , 41 ] platelets [ 42 ] and decrease the risk of coronary heart disease [ 43 ].
Table 7.3 Effects of processing conditions on anthocyanin contents (mg/g DW) of fruits [ 33 ]
Fruits
grower
Fresh
Heating (98 °C,
10 min) Freezing (20 °C) Freeze-drying
Cyd-
3- glu
Cyd-
3- rut
Cyd-
3- glu
Cyd-
3- rut
Cyd-
3- glu
Cyd-
3- rut
Cyd-
3- glu
Cyd-
3- rut
Cherries
Cromwell 207 276 419 475 570 737 158 211
Roxburgh 546 698 470 615 548 706 522 628
Dunedin 46 49 126 172 62 89 22 28
Nectarines
Cromwell 24 25 27 18 33 29 58 7
Roxburgh 16 16 12 12 19 18 71 9
Dunedin 10 15 5 4 9 7 69 7
Peaches
Cromwell 7 7 12 10 17 18 10 12
Roxburgh 16 17 32 27 33 29 6 8
Dunedin 14 15 10 9 14 8 11 15
Plums
Cromwell 51 52 38 40 58 63 14 18
Roxburgh 12 13 37 46 46 59 7 8
Dunedin 18 23 48 60 66 85 19 24
Cyd-3-glu cyaniding-3-glucoside, Cyd-3-rut cyaniding-3-rutinoside
7 The Role of Anthocyanins in Health as Antioxidant, in Bone Health and as Heart…
93
The purposed mechanism of actions include antioxidant effects, modulation of
hormonal and enzyme system, stimulation of the immune system and their inter-
related mechanisms. As mentioned in Chap. 5 , anthocyanins show less bioavailability,
due to their instability under physiological and pH conditions at absorption site in
human body, it was diffi cult to attest their tagged health-promoting effects in animal
models in vivo. Although mechanism of action and various health- promoting effects
need further investigation, anthocyanins certainly have a key role in a healthy and
well-being oriented diet.
7.6 Antioxidant Activity
Anthocyanins exhibit excellent in vitro and in vivo antioxidant activity protecting
human body against oxidative stress by scavenging free radicals and suppressing
lipid peroxidation. Despite various limitations, much progress has been done during
last decade especially in assessing the dynamics of the interplay of ROS and tran-
scription. However the full extent to which anthocyanin is functionally signifi cant
in maintaining the cellular redox homeostasis still remains to be understood. The
core strategy for improving immune suppression and slowing-down the progression
of associated diseases is decreasing the extra production of ROS.
The structure activity relationships of anthocyanins may be considered through
various angles like the position and numbers of hydroxyl and methyl groups in
anthocyanins nucleus. For example the increase antioxidant effect and higher pro-
apoptotic activity in human leukemia cells is considered to be associated with
greater number of hydroxyl group. Similarly the antiproliferative activity is thought
to be more potent with the hydroxyl or methyl groups substitution or placement on
ring B. The phenolic structure of anthocynains imparts antioxidant property to
them. Their antioxidant activity depends on many factors mainly; the number of OH
group; the catechol moiety in B-ring and oxonium ion in C-ring, the methylation
and hydroxylation pattern as well as glycosylation and acylation [ 44 ]. Glycosylation
decreases antioxidant activity as it decreases the capacity of anthocyanin to delocal-
ize electrons [ 45 ]. The contribution of B-ring substituition in antioxidant activity is
OH > OCH3 ≫ H [ 46 ]. The positively charged O-atom in anthocyanin makes it a
potential H-donating antioxidant [ 11 ].
Anthocyanins can express their antioxidant effects directly or indirectly. Directly
they exhibit antioxidant activity by direct free radical scavenging potential due to
electron/hydrogen donating capacity of anthocyanin structure [ 47 , 48 ] which can
bind reactive free radical species. Indirectly, anthocyanins can (i) restore or increase
the activities of glutathione peroxidase or superoxide dismutase thus increasing
glutathione content [ 49 ], (ii) activate genes that code for these enzymes [ 50 ] and
(iii) decrease the generation of oxidative abducts in DNA decreasing the generation
of ROS by inhibiting xanthine oxidase and NADH oxidase or by modifi cation of
arachidonic metabolism and mitochondrial respiration [ 1 , 51 ].
7.6 Antioxidant Activity
94
The anthocyanidins and anthocyanins have shown a higher antioxidant activity
than vitamins C, butylated hydroxyl anisole (BHA), butylated hydroxyl toulene
(BHT), α-tocopherol and Vitamin-E [ 48 , 52 ] by capturing free radicals through the
donation of hydrogen atoms [ 53 , 54 ].
Anthocyanidins have greater antioxidant capacity than anthocyanins i.e. the radi-
cal scavenging activity decreases with glycosylation [ 48 ]. However, Kähkönen and
Heinonen [ 55 ] concluded that in vitro effect of glycosylation on antioxidant activity
is dependent on the environment e.g. anthocyanidin and oxidation models in which
oxidations is occurring [ 56 ].
The antioxidant ability of anthocyanins depends on the chemical structure of a
anthocyanins. It is not necessary that all of them possess similar activities for scaveng-
ing different ROS and reactive nitrogen species (RNS) [ 56 , 57 ] for example the
reactivity against superoxide anion follows the order delphinidine > cyaniding > pelar-
gonidin while pelargonidin is the most effective against the hydroxyl radical [ 58 , 59 ].
Number and position of sugar residues in the anthocyanidin also affects the anti-
oxidant activity of anthocyanins [ 55 , 59 ]. The smaller the number of sugar units at
C3, higher is the antioxidant potential however the effect is method dependent [ 55 ,
60 ]. Pyranoanthocyanins of malvidin, petunidin, pelargonidin and cyanidin exhib-
ited higher superoxide anion radicals (O
2−
) quenching activity but did not quench
hydroxyl radicals [ 61 ], however integration of pyruvic acid into malvidin-3-
monoglucoside and delphinidin-3-monoglucoside decreased signifi cantly the anti-
oxidant activity in aqueous-phase systems as suggested by some reports [ 57 , 60 ].
Greater the number of free OH around the pyrone ring, greater is antioxidant
activity. Anthocyanins with 3′, 4′-dihydroxy groups are good chelating agent for
metal ions [ 62 ]. The chelating capacity also increases at pH 2–4, anthocyanins
exist as fl avylium cations and due to charge distribution, these are vulnerable to
nucleophilic attack on 2 and 4 positions [ 63 ]. Not only the hydroxyl group but the
position and degree of methoxyl groups on pyrone ring infl uenced antioxidant
potential of anthocyanins [ 60 ]. For example cyanidin-3-rutinoside and delphini-
din-3-glucoside have greater antioxidant capacity than malvidin-3-glucoside and
petunidin-3- glucoside [ 55 , 57 ].
The antioxidant potential of berries like red raspberries, black raspberries, straw-
berries and black berries, is directly proportional to the anthocyanins content and
has strong scavenging capacity against chemically generated reactive oxygen spe-
cies [ 64 – 66 ]. Various drinks like whiskeys, sake, Cavas, Chilean Cabernet Sauvignon
red wine, port wine or by-products such as grape pomace and fruit and vegetable
juices have been reported for antioxidant activity due to presence of anthocyanins
[ 67 – 71 ] and a reason behind the French paradox. Red wine fractions having highest
anthocyanins substantially decreased reactive oxygen species (ROS) in human red
blood cells treated with H
2
O
2
as in vitro oxidative model [ 72 ]. Recent days the cus-
tomers expect natural pigment in wine, food and pharmaceutical industry with anti-
oxidant potentials as well [ 73 ].
Wang and Mazza reported that cyaniding, pelargonidin, peonidin, malvidin,
delphinidin, malvidin-3,5-diglucoside and malviding-3-glucoside possess strong inhibi-
tory effects >50 % comparable quercetin on NO production in LPS/IFN-γ- activated
7 The Role of Anthocyanins in Health as Antioxidant, in Bone Health and as Heart…
95
RAW 264.7 macrophage without showing any cytotoxicity at the range of 16–500 μM
[ 34 ]. Anthocyanin-rich berry extracts inhibited NO-production and inhibitory effects are
directly related to the content of total anthocyanins. The antioxidant capacity of serum
increases with the consumption of red wine, strawberries and blueberries the report was
confi rmed by measuring antioxidant capacity as ORAC, TEAC and TRAP [ 74 , 75 ].
Anthocyanins are powerful antioxidants in vitro. Using the oxygen radical absor-
bance capacity (ORAC) assay, antioxidant potential of 14 anthocyanins including
delphinidin, cyanidin, pelargonidin, malvidin, peonidin, and their glycosylated
derivatives was measured in aqueous phase at neutral pH [ 5 ]. Cyanidin-3-glucoside
was found to the highest antioxidant (ORAC value, 3.5 times as potent as Trolox)
and pg exhibited the lowest ORAC value.
In linoleic acid autoxidation, liposome rabbit erythrocyte membrane, and rat
liver microsomal systems, cyanidin-3-glucoside and its aglycone cyanidin were
shown to have similar antioxidant potency as vitamin E (α-tocopherol) [ 76 ].
Cyanidin-3-glucoside diminished powerfully the changes of biomarkers in hepatic
injury in vivo (rat model) hepatic ischemia-reperfusion as an oxidative stress models
thus revealing protective effect of anthocyanins on oxidative stress–induced damage
[ 77 ]. Ramirez-Tortosa et al. found that anthocyanins enhanced plasma antioxidant
potential and reduced the level of 8-oxo-deoxyguanosine and hydroperoxides in
study of 12 week interval on rats, feeding them vitamin E–defi cient diets for followed
by supplementation with purifi ed anthocyanin-rich extracts thus indicating signifi cant
reductions of the vitamin E defi ciency–induced lipid peroxidation and DNA damage
[ 78 ]. The plasma antioxidant status of Dahl–SS rats, measured by a total antioxidant
status (TAS) assay, was restored by a diet supplemented with 1 % freeze-dried whole
tart cherry powder for 90 days [ 79 ]. Various antioxidant assays used for assessment
of antioxidant potential of anthocyanins are shown in Table 7.4 .
Tyrosine nitration activity of anthocyanins declined as: cyanidin-3- rutinoside > mal-
vidin-3-monoglucoside ≈ delphinidin-3- monoglucoside > petunidin- 3-
monoglucoside [ 60 ]. Antioxidant activity is dependent on pH and anthocyanins
isomeric forms [ 77 ]. The peroxynitrite (ONOO
−
) quenching activity of anthocyanins
at pH 7.4 declinedas: cyanidin-3-rutinoside > malvidin-3- monoglucoside ≈ delphinidin-
3-mono-glucoside > petunidin-3-monoglucoside [ 60 ].
7.7 Protection Against Cardiovascular Diseases
The development of cardiovascular disease is mainly due to hypertension, platelet
aggregation, dysfunctioning of vascular endothelium and high plasma LDL choles-
terol. Anthocyanin exerted cardio-protection under ischemia-perfusion condition
besides lowering the oxidative stress in vascular endothelium [ 97 ]. Youdim et al.
(2000) have proved that anthocyanins are incorporated into the cytosol and
membrane of vascular endothelial cells thus protecting against oxidative stress and
protecting endothelial function thereby halting the vascular diseases [ 98 ]. In a study
reporting direct effects of bilberry anthocyanins on whole rat hearts under
7.7 Protection Against Cardiovascular Diseases
96
ischemia- reperfusion situation, the results indicated that perfusion with small
amounts of bilberry anthocyanins (0.01–1 mg/L) substantially reduced ischemia-
perfusion injury by increasing post-ischemic coronary fl ow, decreasing the rate of
lactate dehydrogenase as well as chances and length of reperfusion arrhythmias
[ 99 ]. Anaerobic metabolism can decrease intracellular pH under reperfusion condi-
tions thus affecting radical-quenching activities of anthocyanins [ 100 ]. All these
results suggest that anthocyanin despite their low bioavailability act as cardio-
protectants. In in vivo hamster animal model of ischemia-reperfusion, bilberry
anthocyanin decreased micro-vascualr injuries by preserving endothelium and
enhanced capillary perfusion [ 101 ]. In 8 week dietary ingestion of plant-derived
anthocyanin in rats, made myocardium less vulnerable to ischemia-reperfusion
impairments in vivo as well as ex vivo [ 49 ]. Figure 7.2 indicates the relationship
between consumption of anthocyanins and their effects on cardiovascular health.
As mentioned above, over production of free radicals and reactive oxygen spe-
cies (ROS) are the key elements that led to activation of pro- infl ammatory media-
tors that ultimately led to cardiovascular disorders like ischemic heart problem,
cardiomyopathy and coronary heart problems. Anthocyanins are most consumed
subgroup of fl avonoids that control/reduce this heart associated chronic abnormali-
ties via antioxidant, anti-infl ammatory and free radical scavenging mechanisms.
Table 7.4 Various antioxidant assays reported for anthocyanins
Name of antioxidant assay Reference
Oxygen radical absorbance capacity (ORAC) [
3 , 5 , 65 , 74 , 75 ,
80 , 81 ]
Hydrogen transfer-based assay [
3 , 5 , 65 , 80 , 82 ]
Ferric reducing antioxidant potential (FRAP) [
83 – 88 ]
Trolox equivalent antioxidant capacity (TEAC) [
74 , 75 , 81 ,
83 – 88 ]
2,2-diphenyl-1-picrylhydrazyl (DPPH) free radical scavenging activity [
83 – 88 ]
Electron transfer-based assays [
83 – 88 ]
Superoxide scavenging assay [
89 ]
Peroxynitrite (ONOO−) scavenging activity [
90 ]
Inhibition of human low-density lipoprotein [
64 ]
Inhibition of lipid peroxidation [
90 ]
Induction of antioxidant enzymes assay e.g. gluthatione- S -transferase
(GST), gluthationereductase (GR), gluthationeperoxidise (GPx) and
superoxide dismutase
[
91 , 92 ]
Heavy metal binding assay e.g. iron, zinc and copper [
93 ]
NO assay [
94 ]
TRAP [
74 , 75 , 81 ]
Human red blood cells treated with H
2
O
2
as in vitro oxidative model [ 95 ]
Hepatic ischemia-reperfusion as an oxidative stress model [
77 ]
Vitamin E defi ciency–induced lipid peroxidation [
78 ]
Serum antioxidant capacity (SAOC) [
96 ]
Total antioxidant status (TAS) assay [
79 ]
7 The Role of Anthocyanins in Health as Antioxidant, in Bone Health and as Heart…
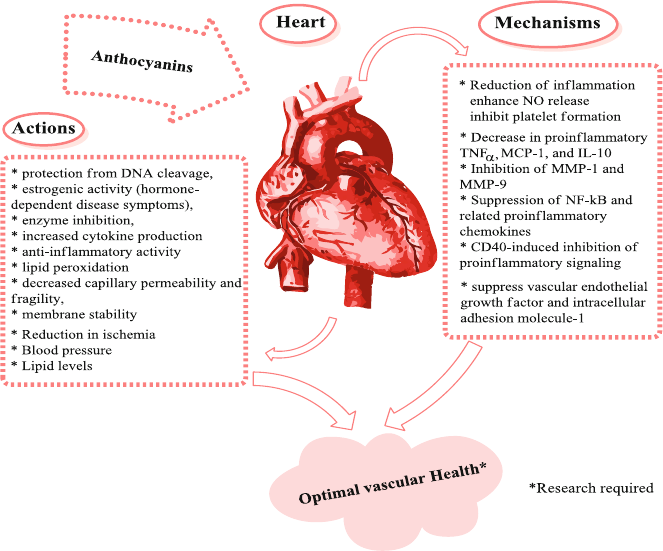
97
Despite the high fat intake there is low incidence of cardiovascular disease amongst
the French people (French paradox) which was supposed to be due to the daily con-
sumption of red wine, rich in polyphenols like anthocyanins [ 43 , 103 ]. Fresh,
freeze-dried and purifi ed extracts and juice of chokeberries, cranberries, blueber-
ries, and strawberries anthocyanins have verifi ed signifi cant step up in low density
lipoprotein (LDL) oxidation, lipid peroxidation, dyslipidemia, glucose metabolism
and total plasma antioxidant status in human blood [ 104 , 105 ].
Mink et al. (2007) reported in a specifi c epidemiological study that strawberry
and blueberry intake caused reduction of mortality due cardiovascular diseases in
postmenopausal woman [ 106 ]. The effect was considered to be due to presence of
anthocyanins, a question was raised that other fl avonoids may also contribute but
later in another epidemiological study, Cassidy et al. reported that high consump-
tion of anthocyanins can decrease MI risk in predominantly young women in a
comparison of those with lower intake. They confi rmed that other fl avonoids are not
associated with such effects in their study [ 107 ] however further details trials are
required to note the impact of anthocyanins enriched food.
According to several epidemiological studies, coronary heart disease mortality
may be reduced by reasonable intake of red wine [ 108 ]. The reduction in mortality
due to coronary heart disease may be due to reduction in platelet coagulation [ 43 ],
Fig. 7.2 Anthocyanins and cardiovascular health (actions and mechanisms) [ 102 ]
7.7 Protection Against Cardiovascular Diseases
98
increase in circulatory high-density lipoprotein (HDL) [ 108 ], free radical scaveng-
ing, modulation of eicosanoid metabolism and inhibition of lipoprotein oxidation
[ 109 ]. The last three mechanisms are also considered to be responsible for reduction
of atherosclerosis [ 75 ]. Anthocyanins are responsible for reduction in coronary
heart diseases and other health benefi cial effects so complete metabolomic profi le is
required in properly designed long term intervention studies. This area has much
space to be investigated [ 75 ].
Daily intake of concentrated red grape juice (@125 mL) increase serum anti-
oxidant potential thus reducing susceptibility of LDL to oxidation. Thus red grape
juice was proposed to possess benefi cial effects equal to red wine [ 110 ]. Similarly
other anthocyanins rich foods e.g. black currant consumption cause a rapid growth
in plasma antioxidant capacity until 2 h [ 111 ]. Abuja et al. investigated that spray-
dried elderberry juice comprising greater anthocyanins content has shielding
effect on human LDL in vitro [ 112 ]. Tsuda et al. (1996) demonstrate that purifi ed
anthocyanins like pelagonidin-3-glucoside, cyanidin-3-glucoside, delphenidin- 3-
glucoside and their aglycones in a UV-induced lipid peroxidation model, showed
strong inhibition of lipid peroxidation [ 58 ]. Even anthocyanin colonic metabolite,
i.e. protocatechuic acid improved atherosclerosis progression by acting as
anti- infl ammatory agent and also exhibited antiplatelet activity [ 113 , 114 ].
7.8 Anthocyanins and Bone Health
Bone health is one of the prime concerns in human health specially aged people;
one of the most common occurring bone diseases is osteoporosis. Osteoporosis is
characterized by decrease in bone mineral content including decrease in calcium
content; age related factors like hormonal imbalance, chronic infl ammation and
increase in oxidative stress are the main causes [ 115 ]. All these factors are linked to
reactive oxygen species and to prevent bone loss, direct or indirect control of reac-
tive oxygen species is required. Anthocyanins containing foods are thought to have
important role in the prevention of osteoporosis. Different studies reported the con-
fi rmed the role of anthocyanins in bone diseases prevention (Fig. 7.3 ).
Kaume et al. (2015) carried out a study in 9 months old female Sprague–Dawley
rats, it was confi rmed that feeding blackberries to rats prevent bone desorption
especially in ovariectomized rat models, the author observed the effect in dose
dependent manner, cyanidin 3- O - β - D -glucoside may be responsible for the bone
loss preventive effect [ 116 ]. Women who consumed fruit in greater quantity, in
childhood have high bone mineral density compared to normal, similar is the case
with all women and men who are consumer of fruit, vegetables and grains [ 115 ,
117 ]. Welch et al. (2013) carried out an observational study in women that were
habitual to fl avonoids and anthocyanins intake, they observed increased bone min-
eral density [ 118 ]. Similarly the decrease risk of fracture was observed by
Langestmo et al. (2011) in individuals that were frequents consumers of fruit,
vegetables and grains [ 119 ].
7 The Role of Anthocyanins in Health as Antioxidant, in Bone Health and as Heart…
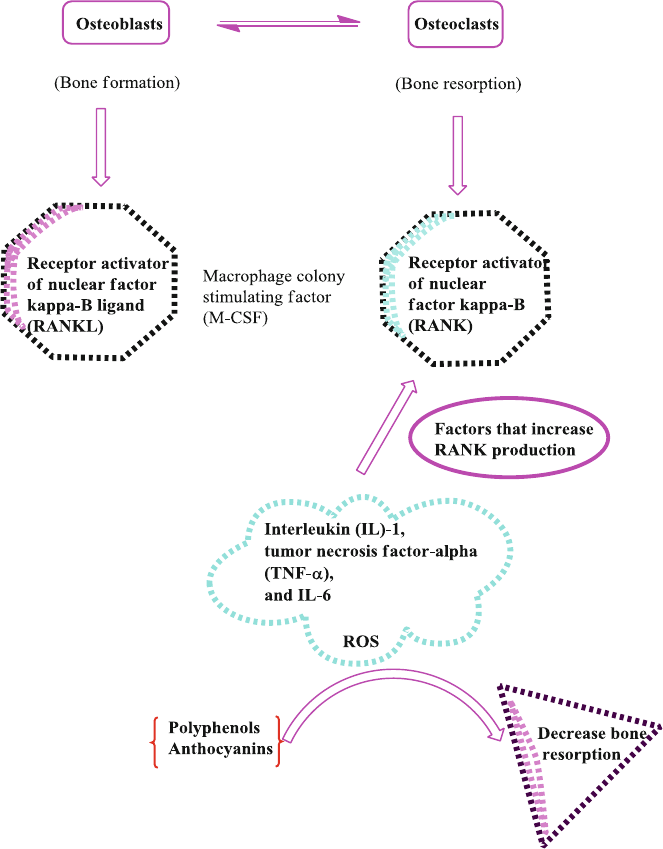
99
Tanabe et al. (2011) applied cranberry extract on human bone marrow cells
(pre- osteoclastic) at a dose of 10, 25, 50, 100 μg/mL for 4 days duration, the rate of
bone degradation by inhibition of RANKL dependent osteoclasts decreased [ 120 ].
500 ng/mL blueberry extract for 72 h duration increased proliferation of human
bone marrow cells (CD34+ or CD133) while decreased tartrate resistant acid phos-
phatase (TRAP) staining and RANKL dependent osteoclast numbers [ 121 ]. Six
months old SHAM and OVX (ovariectomized) female rats, were used in this study,
Fig. 7.3 Anthocyanins osteoporosis control mechanism
7.8 Anthocyanins and Bone Health
100
they were treated in three groups as (i) SHAM Control (ii) OVX-Control and (iii)
OVX + 5 % blueberry for 100 day duration, it was found that OVX + 5 % blueberry
group increased whole body bone mineral density (BMD) and serum alkaline phos-
phatase (ALP) [ 122 ].
Chen et al. (2010) and Zhang et al. (2011) studied the affect of blueberry enriched
diet in Sprague–Dawley rats and it was found that bone mass and bone mineral
density increases, Early blueberry supplementation prevented osteoblast senescence
and adult bone loss [ 123 , 124 ].
Berries are rich source of anthocyanins so the bone loss preventive effect are
considered to associated with the antioxidant, and anti-infl ammatory effect of
anthocyanins however as berries also contain vitamin C so it’s could not be excluded.
Dou et al. (2014) studied the effect of cyanidin on bone for the fi rst time, they
used RAW264.7 cells and primary bone monocytes in their study design, they found
that cyanidin has dual effect on RANKL induced osteoclastogenesis, dose less than
1 μg has promoting while greater than 10 μg has inhibitory effect. Fusogenic and
osteoclast related genes are regulated by cyanidin by the same dual pattern thus
osteoclast related bone disorders may be controlled with the use of cyanidin [ 125 ].
Related group members of anthocyanins delphinidins have been shown to support
bone mineralization by suppressing the differentiation and function of osteoclasts,
the cells resorbing bone, this statement is supported by study in ovariectomized
mice model through oral administration of delphinidin [ 126 , 127 ]. These investiga-
tions suggest a potential benefi t from regular consumption of anthocyanins rich
sources for preventing bone demineralization at higher age.
7.9 Conclusions
Despite all above mentioned effects, it is occasionally diffi cult to discriminate the
effects of anthocyanins from other constituents or even isolation of effect of a
single anthocyanin molecule from total pool. Further the doses mentioned in lit-
erature for animal models are sometimes far from those compatible with realistic
values for human body due to bioavailability differences in both. There are entic-
ing glimpses of health-promoting effects however many questions remain unan-
swered since most of these studies are in vitro. There is need of balanced,
data-driven answers for questions raised by nutritionists and health professionals
regarding the totality of evidence rather than fi ndings of any single study. There is
still unceratininity about some mechanisms. There is plenty of information about
role of anthocyanins in prevention of major diseases however many tropical and
emerging diseases remain un- attentioned. Considering complex biological
functions and their effi cacy in prevention of various chronic human diseases,
combination therapies with other natural antioxidants can be a useful approach for
various diseases.
7 The Role of Anthocyanins in Health as Antioxidant, in Bone Health and as Heart…
101
References
1. Pojer, E., Mattivi, F., Johnson, D., & Stockley, C. S. (2013). The case for anthocyanin con-
sumption to promote human health: A review. Comprehensive Reviews in Food Science and
Food Safety, 12 (5), 483–508.
2. Timberlake, C. (1988). The biological properties of anthocyanin compounds. NATCOL
Quarterly Bulletin, 1 , 4–15.
3. Prior, R. L., Cao, G., Martin, A., Sofi c, E., McEwen, J., O’Brien, C., et al. (1998). Antioxidant
capacity as infl uenced by total phenolic and anthocyanin content, maturity, and variety of
Vaccinium species. Journal of Agricultural and Food Chemistry, 46 (7), 2686–2693.
4. Degenhardt, A., Knapp, H., & Winterhalter, P. (2000). Separation and purifi cation of antho-
cyanins by high-speed countercurrent chromatography and screening for antioxidant activity.
Journal of Agricultural and Food Chemistry, 48 (2), 338–343.
5. Wang, H., Cao, G., & Prior, R. L. (1997). Oxygen radical absorbing capacity of anthocyanins.
Journal of Agricultural and Food Chemistry, 45 (2), 304–309.
6. Lietti, A., Cristoni, A., & Picci, M. (1976). Studies on Vaccinium myrtillus anthocyanosides.
I. Vasoprotective and anti-infl amatory activity. Arzneimittel-Forschung, 26 (5), 829–832.
7. Morazzoni, P., & Magistretti, M. (1986). Effects of Vaccinium myrtillus anthocyanosides on
prostacyclin-like activity in rat arterial tissue. Fitoterapia, 57 , 11–14.
8. Kamei, H., Kojima, T., Hasegawa, M., Koide, T., Umeda, T., Yukawa, T., et al. (1995).
Suppression of tumor cell growth by anthocyanins in vitro. Cancer Investigation, 13 (6),
590–594.
9. Karaivanova, M., Drenska, D., & Ovcharov, R. (1990). A modifi cation of the toxic effects of
platinum complexes with antocyans. Eksperimentalna Meditsina i Morfologiia, 29 (2), 19.
10. Akhmadieva, A. K., Zaichkina, S. I., Ruzieva, R. K., & Ganassi, E. E. (1993). The protective
action of a natural preparation of anthocyan (pelargonidin-3, 5-diglucoside). Radio Biologica,
33 (3), 433.
11. Kong, J.-M., Chia, L. S., Goh, N. K., Chia, T. F., & Brouillard, R. (2003). Analysis and bio-
logical activities of anthocyanins. Phytochemistry, 64 (5), 923–933.
12. Valls, J., Millán, S., Martí, M. P., Borràs, E., & Arola, L. (2009). Advanced separation meth-
ods of food anthocyanins, isofl avones and fl avanols. Journal of Chromatography A, 1216 (43),
7143–7172.
13. Cardile, V., Frasca, G., Rizza, L., Rapisarda, P., & Bonina, F. (2010). Anti-infl ammatory
effects of a red orange extract in human keratinocytes treated with interferon gamma and
histamine. Phytotherapy Research, 24 (3), 414–418.
14. Lorencini, M., Brohem, C. A., Dieamant, G. C., Zanchin, N. I., & Maibach, H. I. (2014).
Active ingredients against human epidermal aging. Ageing Research Reviews, 15 , 100–115.
15. Mantena, S. K., & Katiyar, S. K. (2006). Grape seed proanthocyanidins inhibit UV-radiation-
induced oxidative stress and activation of MAPK and NF-κB signaling in human epidermal
keratinocytes. Free Radical Biology and Medicine, 40 (9), 1603–1614.
16. Scalbert, A., & Williamson, G. (2000). Dietary intake and bioavailability of polyphenols.
Journal of Nutrition, 130 (8), 2073S–2085S.
17. Prior, R. L., & Wu, X. (2006). Anthocyanins: Structural characteristics that result in unique
metabolic patterns and biological activities. Free Radical Research, 40 (10), 1014–1028.
18. Wang, S. Y., & Jiao, H. (2000). Scavenging capacity of berry crops on superoxide radicals,
hydrogen peroxide, hydroxyl radicals, and singlet oxygen. Journal of Agricultural and Food
Chemistry, 48 (11), 5677–5684.
19. Shih, P.-H., & Yen, G.-C. (2007). Differential expressions of antioxidant status in aging rats:
The role of transcriptional factor Nrf2 and MAPK signaling pathway. Biogerontology, 8 (2),
71–80.
20. Brouillard, R. (1982). Chemical structure of anthocyanins (Vol. 1). New York: Academic.
21. Markakis, P. (2012). Anthocyanins as food colors . New York: Elsevier.
22. Morazzoni, P., & Bombardelli, E. (1996). Vaccinium myrtillus L. Fitoterapia, 67 (1), 3–29.
References
102
23. He, J., & Giusti, M. M. (2010). Anthocyanins: Natural colorants with health-promoting prop-
erties. Annual Review of Food Science and Technology, 1 , 163–187.
24. Brouillard, R., & Markakis, P. (1982). Anthocyanins as food colors . New York: Academic.
25. Eder, R., & Nollet, L. (2000). Pigments. In M. L. L. Nollet (Ed.), Food analysis by HPLC (pp.
825–880). New York: Marcer Dekker.
26. Organization, W. H. (1982). Anthocyanins. Toxicological Evaluation of Food Additives:
Technical Report Series, 17 , 42–49.
27. Clifford, M. N. (2000). Anthocyanins-nature, occurrence and dietary burden. Journal of the
Science of Food and Agriculture, 80 (7), 1063–1072.
28. EFSA Panel on Food Additives and Nutrient Sources added to Food. (2013). Statement on
two reports published after the closing date of the public consultation of the draft Scientifi c
Opinion on the re-evaluation of aspartame (E 951) as a food additive. European Food Safety
Authority Journal, 11 (12), 3504.
29. Brown, J. P., & Dietrich, P. S. (1979). Mutagenicity of plant fl avonols in the Salmonella/
mammalian microsome test: Activation of fl avonol glycosides by mixed glycosidases from
rat cecal bacteria and other sources. Mutation Research, 66 (3), 223–240.
30. Macgregor, J. T., & Jurd, L. (1978). Mutagenicity of plant fl avonoids: Structural require-
ments for mutagenic activity in Salmonella typhimurium. Mutation Research, 54 (3),
297–309.
31. Haveland-Smith, R. (1981). Evaluation of the genotoxicity of some natural food colours
using bacterial assays. Mutation Research Letters, 91 (4), 285–290.
32. Cox, G., Rucci, G., & Babish, J. (1978). 90-day subacute dietary toxicity study of 78-002-2
in Sprague-Dawley rats. Unpublished report submitted to the Flavor and Extract Manufacturers
Association, Washington, DC, USA, by Food and Drug Research Laboratories, Inc. Submitted
to WHO by the International Organization of the Flavor Industry, Brussels, Belgium.
33. Leong, S. Y., & Oey, I. (2012). Effects of processing on anthocyanins, carotenoids and vita-
min C in summer fruits and vegetables. Food Chemistry, 133 (4), 1577–1587.
34. Wang, J., & Mazza, G. (2002). Effects of anthocyanins and other phenolic compounds on the
production of tumor necrosis factor α in LPS/IFN-γ-activated RAW 264.7 macrophages.
Journal of Agricultural and Food Chemistry, 50 (15), 4183–4189.
35. Pisha, E., & Pezzuto, J. (1994). Fruits and vegetables containing compounds that demonstrate
pharmacological activity in humans. Economic and Medicinal Plant Research, 6 , 189–233.
36. Katsube, N., Iwashita, K., Tsushida, T., Yamaki, K., & Kobori, M. (2003). Induction of apop-
tosis in cancer cells by bilberry (Vaccinium myrtillus) and the anthocyanins. Journal of
Agricultural and Food Chemistry, 51 (1), 68–75.
37. Matsumoto, H., Nakamura, Y., Iida, H., Ito, K., & Ohguro, H. (2006). Comparative assess-
ment of distribution of blackcurrant anthocyanins in rabbit and rat ocular tissues. Experimental
Eye Research, 83 (2), 348–356.
38. Youdim, K., Shukitt-Hale, B., MacKinnon, S., Kalt, W., & Joseph, J. A. (2000). Polyphenolics
enhance red blood cell resistance to oxidative stress: In vitro and in vivo. Biochimica et
Biophysica Acta, 1523 (1), 117–122.
39. Galli, R. L., Shukitt-Hale, B., Youdim, K. A., & Joseph, J. A. (2002). Fruit polyphenolics and
brain aging: nutritional interventions targeting age-related neuronal and behavioral defi cits.
Annals of the New York Academy of Science, 959 (1), 128–132.
40. Andriambeloson, E., Magnier, C., Haan-Archipoff, G., Lobstein, A., Anton, R., Beretz, A., et
al. (1998). Natural dietary polyphenolic compounds cause endothelium-dependent vasore-
laxation in rat thoracic aorta. Journal of Nutrition, 128 (12), 2324–2333.
41. Martin, P. D., Warwick, M. J., Dane, A. L., Brindley, C., & Short, T. (2003). Absolute oral
bioavailability of rosuvastatin in healthy white adult male volunteers. Clinical Therapeutics,
25 (10), 2553–2563.
42. Demrow, H. S., Slane, P. R., & Folts, J. D. (1995). Administration of wine and grape juice
inhibits in vivo platelet activity and thrombosis in stenosed canine coronary arteries.
Circulation, 91 (4), 1182–1188.
7 The Role of Anthocyanins in Health as Antioxidant, in Bone Health and as Heart…
103
43. Renaud, S. D., & de Lorgeril, M. (1992). Wine, alcohol, platelets, and the French paradox for
coronary heart disease. Lancet, 339 (8808), 1523–1526.
44. Yang, M., Koo, S. I., Song, W. O., & Chun, O. K. (2011). Food matrix affecting anthocyanin
bioavailability: Review. Current Medicinal Chemistry, 18 (2), 291–300.
45. Wang, L.-S., & Stoner, G. D. (2008). Anthocyanins and their role in cancer prevention.
Cancer Letters, 269 (2), 281–290.
46. Rossetto, M., Vanzani, P., Lunelli, M., Scarpa, M., Mattivi, F., & Rigo, A. (2007). Peroxyl
radical trapping activity of anthocyanins and generation of free radical intermediates. Free
Radical Research, 41 (7), 854–859.
47. Borkowski, T., Szymusiak, H., Gliszczyńska-Rwigło, A., Rietjens, I. M., & Tyrakowska, B.
(2005). Radical scavenging capacity of wine anthocyanins is strongly pH-dependent. Journal
of Agricultural and Food Chemistry, 53 (14), 5526–5534.
48. Fukumoto, L., & Mazza, G. (2000). Assessing antioxidant and prooxidant activities of phe-
nolic compounds. Journal of Agricultural and Food Chemistry, 48 (8), 3597–3604.
49. Toufektsian, M.-C., de Lorgeril, M., Nagy, N., Salen, P., Donati, M. B., Giordano, L., et al.
(2008). Chronic dietary intake of plant-derived anthocyanins protects the rat heart against
ischemia-reperfusion injury. Journal of Nutrition, 138 (4), 747–752.
50. Shih, P.-H., Yeh, C.-T., & Yen, G.-C. (2005). Effects of anthocyanidin on the inhibition of
proliferation and induction of apoptosis in human gastric adenocarcinoma cells. Food and
Chemical Toxicology, 43 (10), 1557–1566.
51. Steffen, Y., Gruber, C., Schewe, T., & Sies, H. (2008). Mono-O-methylated fl avanols and
other fl avonoids as inhibitors of endothelial NADPH oxidase. Archives of Biochemistry and
Biophysics, 469 (2), 209–219.
52. Bagchi, D., Garg, A., Krohn, R. L., Bagchi, M., Bagchi, D. J., Balmoori, J., et al. (1998).
Protective effects of grape seed proanthocyanidins and selected antioxidants against TPA-
induced hepatic and brain lipid peroxidation and DNA fragmentation, and peritoneal macro-
phage activation in mice. General Pharmacology, 30 (5), 771–776.
53. Chen, Z. Y., Chan, P. T., Ho, K. Y., Fung, K. P., & Wang, J. (1996). Antioxidant activity of
natural fl avonoids is governed by number and location of their aromatic hydroxyl groups.
Chemistry and Physics of Lipids, 79 (2), 157–163.
54. Rice-Evans, C. A., Miller, N. J., & Paganga, G. (1996). Structure-antioxidant activity rela-
tionships of fl avonoids and phenolic acids. Free Radical Biology and Medicine, 20 (7),
933–956.
55. Kähkönen, M. P., & Heinonen, M. (2003). Antioxidant activity of anthocyanins and their
aglycons. Journal of Agricultural and Food Chemistry, 51 (3), 628–633.
56. Kay, C. D., Mazza, G., Holub, B. J., & Wang, J. (2004). Anthocyanin metabolites in human
urine and serum. British Journal of Nutrition, 91 (06), 933–942.
57. Miguel, M. (2011). Anthocyanins: Antioxidant and/or anti-infl ammatory activities .
58. Tsuda, T., Shiga, K., Ohshima, K., Kawakishi, S., & Osawa, T. (1996). Inhibition of lipid
peroxidation and the active oxygen radical scavenging effect of anthocyanin pigments iso-
lated from Phaseolus vulgaris L. Biochemical Pharmacology, 52 (7), 1033–1039.
59. Antal, D.-S., Garban, G., & Garban, Z. (2003). The anthocyans: Biologicallyactive sub-
stances of food and pharmaceutic interest. Annals of the University Dunarea de Jos of Galati,
Food Technology, 6 , 106–115.
60. Muselík, J., García-Alonso, M., Martín-López, M. P., Žemlička, M., & Rivas-Gonzalo, J. C.
(2007). Measurement of antioxidant activity of wine catechins, procyanidins, anthocyanins
and pyranoanthocyanins. International Journal of Molecular Science, 8 (8), 797–809.
61. Garcia Alonso, M., Rimbach, G., Sasai, M., Nakahara, M., Matsugo, S., Uchida, Y., et al.
(2005). Electron spin resonance spectroscopy studies on the free radical scavenging activity
of wine anthocyanins and pyranoanthocyanins. Molecular Nutrition and Food Research,
49 (12), 1112–1119.
62. Sarma, A. D., Sreelakshmi, Y., & Sharma, R. (1997). Antioxidant ability of anthocyanins
against ascorbic acid oxidation. Phytochemistry, 45 (4), 671–674.
References
104
63. Bkowska-Barczak, A. (2005). Acylated anthocyanins as stable, natural food colorants – A
review. Polish Journal of Food and Nutrition Sciences, 14 (2), 107–116.
64. Heinonen, I. M., Meyer, A. S., & Frankel, E. N. (1998). Antioxidant activity of berry pheno-
lics on human low-density lipoprotein and liposome oxidation. Journal of Agricultural and
Food Chemistry, 46 (10), 4107–4112.
65. Wang, S. Y., & Lin, H.-S. (2000). Antioxidant activity in fruits and leaves of blackberry,
raspberry, and strawberry varies with cultivar and developmental stage. Journal of Agricultural
and Food Chemistry, 48 (2), 140–146.
66. Castañeda-Ovando, A., de Lourdes, P. H. M., Elena, P. H., José, A. R., & Carlos, A. G. V.
(2009). Chemical studies of anthocyanins: A review. Food Chemistry, 113 (4), 859–871.
67. McPhail, D. B., Peter, T. G., Garry, G. D., Gordon, M. S., & Kenneth, R. (1999). Assessment
of the antioxidant potential of Scotch whiskeys by electron spin resonance spectroscopy:
Relationship to hydroxyl-containing aromatic components. Journal of Agricultural and Food
Chemistry, 47 (5), 1937–1941.
68. Kitagaki, H., & Tsugawa, M. (1999). 1,1-Diphenyl-2-picrylhydrazyl radical (DPPH) scav-
enging ability of sake during storage. Journal of Bioscience and Bioengineering, 87 (3),
328–332.
69. Satué-Gracia, M. T., Andrés-Lacueva, C., Lamuela-Raventós, R. M., & Frankel, E. N. (1999).
Spanish sparkling wines (cavas) as inhibitors of in vitro human low-density lipoprotein oxi-
dation. Journal of Agricultural and Food Chemistry, 47 (6), 2198–2202.
70. Aguirre, M. J., Chen, Y. Y., Mauricio, I., Betty, M., Leonora, M., & Simonet, T. (2010).
Electrochemical behaviour and antioxidant capacity of anthocyanins from Chilean red wine,
grape and raspberry. Food Chemistry, 121 (1), 44–48.
71. Oliveira, B. F., Nogueira-Machado, J. A., & Chaves, M. M. (2010). The role of oxidative
stress in the aging process. Scientifi c World Journal, 10 , 1121–1128.
72. Tedesco, I., Luigi Russo, G., Nazzaro, F., Russo, M., & Palumbo, R. (2001). Antioxidant
effect of red wine anthocyanins in normal and catalase-inactive human erythrocytes. Journal
of Nutrition and Biochemistry, 12 (9), 505–511.
73. Li, X., Ma, H., Huang, H., Li, D., & Yao, S. (2013). Natural anthocyanins from phytore-
sources and their chemical researches. Natural Product Research, 27 (4-5), 456–469.
74. Mazza, G., Kay, C. D., Cottrell, T., & Holub, B. J. (2002). Absorption of anthocyanins from
blueberries and serum antioxidant status in human subjects. Journal of Agricultural and Food
Chemistry, 50 (26), 7731–7737.
75. Mazza, G. (2007). Anthocyanins and heart health. Annali dell’Istituto Superiore di Sanità,
43 (4), 369.
76. Tsuda, T., Mie, W., Katsumi, O., Seiji, N., Sang-Won, C., Shunro, K., et al. (1994).
Antioxidative activity of the anthocyanin pigments cyanidin 3-O-β-D-glucoside and cyanidin.
Journal of Agricultural and Food Chemistry, 42 (11), 2407–2410.
77. Tsuda, T., Horio, F., & Osawa, T. (2000). The role of anthocyanins as an antioxidant under
oxidative stress in rats. Biofactors, 13 (14), 133–139.
78. Ramirez-Tortosa, C., Andersen, Ø. M., Cabrita, L., Gardner, P. T., Morrice, P. C., Wood, S.
G., et al. (2001). Anthocyanin-rich extract decreases indices of lipid peroxidation and DNA
damage in vitamin E-depleted rats. Free Radical Biology and Medicine, 31 (9), 1033–1037.
79. Seymour, E. M., Singer, A. A., Kirakosyan, A., Urcuyo-Llanes, D. E., Kaufman, P. B., &
Bolling, S. F. (2008). Altered hyperlipidemia, hepatic steatosis, and hepatic peroxisome pro-
liferator-activated receptors in rats with intake of tart cherry. Journal of Medicinal Food,
11 (2), 252–259.
80. Steed, L., & Truong, V. D. (2008). Anthocyanin content, antioxidant activity, and selected
physical properties of fl owable purple‐fl eshed sweet potato purees. Journal of Food Science,
73 (5), S215–S221.
81. Cao, G., Russell, R. M., Lischner, N., & Prior, R. L. (1998). Serum antioxidant capacity is
increased by consumption of strawberries, spinach, red wine or vitamin C in elderly women.
Journal of Nutrition, 128 (12), 2383–2390.
7 The Role of Anthocyanins in Health as Antioxidant, in Bone Health and as Heart…
105
82. Zheng, W., & Wang, S. Y. (2003). Oxygen radical absorbing capacity of phenolics in blueber-
ries, cranberries, chokeberries, and lingonberries. Journal of Agricultural and Food
Chemistry, 51 (2), 502–509.
83. Moyer, R. A., Hummer, K. E., Finn, C. E., Frei, B., & Wrolstad, R. E. (2002). Anthocyanins,
phenolics, and antioxidant capacity in diverse small fruits: Vaccinium, Rubus, and Ribes.
Journal of Agricultural and Food Chemistry, 50 (3), 519–525.
84. Lee, J., Koo, N., & Min, D. (2004). Reactive oxygen species, aging, and antioxidative nutra-
ceuticals. Comprehensive Reviews in Food Science and Food Safety, 3 (1), 21–33.
85. Einbond, L. S., Einbond, K. A., Reynertson, X. D. L., Margaret, J. B., & Edward, J. K.
(2004). Anthocyanin antioxidants from edible fruits. Food Chemistry, 84 (1), 23–28.
86. Nakajima, J.-I., Tanaka, I., Seo, S., Yamazaki, M., & Saito, K. (2004). LC/PDA/ESI-MS
profi ling and radical scavenging activity of anthocyanins in various berries. BioMed Research
International, 2004 (5), 241–247.
87. Solomon, A., Golubowicz, S., Yablowicz, Z., Grossman, S., Bergman, M., Gottlieb, H. E., et
al. (2006). Antioxidant activities and anthocyanin content of fresh fruits of common fi g
(Ficus carica L.). Journal of Agricultural and Food Chemistry, 54 (20), 7717–7723.
88. Koca, I., & Karadeniz, B. (2009). Antioxidant properties of blackberry and blueberry fruits
grown in the Black Sea Region of Turkey. Scientia Horticulturae, 121 (4), 447–450.
89. Costantino, L., Albasini, A., Rastelli, G., & Benvenuti, S. (1992). Activity of polyphenolic
crude extracts as scavengers of superoxide radicals and inhibitors of xanthine oxidase. Planta
Medica, 58 (4), 342–344.
90. Muselík, J., García-Alonso, M., Martín-López, M. P., Žemlička, M., & Rivas-Gonzalo, J. C.
(2007). Measurement of antioxidant activity of wine catechins, procyanidins, anthocyanins
and pyranoanthocyanins. International Journal of Molecular Sciences, 8 (8), 797–809.
91. Fiander, H., & Schneider, H. (2000). Dietary ortho phenols that induce glutathione
S-transferase and increase the resistance of cells to hydrogen peroxide are potential cancer
chemopreventives that act by two mechanisms: The alleviation of oxidative stress and the
detoxifi cation of mutagenic xenobiotics. Cancer Letters, 156 (2), 117–124.
92. Turner, M. K. (2009). Anthocyanins increase antioxidant enzyme activity in HT-29 adenocar-
cinoma cells . MS Thesis, University of Georgia.
93. Havsteen, B. (1983). Flavonoids, a class of natural products of high pharmacological potency.
Biochemical Pharmacology, 32 (7), 1141–1148.
94. Wang, J., & Mazza, G. (2002). Inhibitory effects of anthocyanins and other phenolic com-
pounds on nitric oxide production in LPS/IFN-γ-activated RAW 264.7 macrophages. Journal
of Agricultural and Food Chemistry, 50 (4), 850–857.
95. Tedesco, I., Luigi Russo, G., Nazzaro, F., Russo, M., & Palumbo, R. (2001). Antioxidant
effect of red wine anthocyanins in normal and catalase-inactive human erythrocytes. Journal
of Nutritional Biochemistry, 12 (9), 505–511.
96. Whitehead, T. P., Robinson, D., Allaway, S., Syms, J., & Hale, A. (1995). Effect of red wine
ingestion on the antioxidant capacity of serum. Clinical Chemistry, 41 (1), 32–35.
97. Ziberna, L., Tramer, F., Moze, S., Vrhovsek, U., Mattivi, F., & Passamonti, S. (2012).
Transport and bioactivity of cyanidin 3-glucoside into the vascular endothelium. Free Radical
Biology and Medicine, 52 (9), 1750–1759.
98. Youdim, K. A., Martin, A., & Joseph, J. A. (2000). Incorporation of the elderberry anthocya-
nins by endothelial cells increases protection against oxidative stress. Free Radical Biology
and Medicine, 29 (1), 51–60.
99. Halliwell, B. (2008). Are polyphenols antioxidants or pro-oxidants? What do we learn from
cell culture and in vivo studies? Archives of Biochemistry and Biophysics, 476 (2), 107–112.
100. Cvorovic, J., Tramer, F., Granzotto, M., Candussio, L., Decorti, G., & Passamonti, S. (2010).
Oxidative stress-based cytotoxicity of delphinidin and cyanidin in colon cancer cells. Archives
of Biochemistry and Biophysics, 501 (1), 151–157.
101. Bertuglia, S., Malandrino, S., & Colantuoni, A. (1995). Effect of Vaccinium myrtillus antho-
cyanosides on ischaemia reperfusion injury in hamster cheek pouch microcirculation.
Pharmacological Research, 31 (3), 183–187.
References
106
102. Wallace, T. C. (2011). Anthocyanins in cardiovascular disease. Advances in Nutrition, 2 (1), 1–7.
103. Kanner, J., Edwin, F., Rina, G., Bruce, G., & John, E. K. (1994). Natural antioxidants in
grapes and wines. Journal of Agricultural and Food Chemistry, 42 (1), 64–69.
104. Basu, A., Rhone, M., & Lyons, T. J. (2010). Berries: Emerging impact on cardiovascular
health. Nutrition Reviews, 68 (3), 168–177.
105. Rodriguez-Mateos, A., Ishisaka, A., Mawatari, K., Vidal-Diez, A., Spencer, J. P., & Terao, J.
(2013). Blueberry intervention improves vascular reactivity and lowers blood pressure in
high-fat-, high-cholesterol-fed rats. British Journal of Nutrition, 109 (10), 1746–1754.
106. Mink, P. J., Scrafford, C. G., Barraj, L. M., Harnack, L., Hong, C. P., Nettleton, J. A., et al.
(2007). Flavonoid intake and cardiovascular disease mortality: A prospective study in post-
menopausal women. American Journal of Clinical Nutrition, 85 (3), 895–909.
107. Cassidy, A., Mukamal, K. J., Liu, L., Franz, M., Eliassen, A. H., & Rimm, E. B. (2013). High
anthocyanin intake is associated with a reduced risk of myocardial infarction in young and
middle-aged women. Circulation, 127 (2), 188–196.
108. Klatsky, A. L., Tekawa, I., Armstrong, M. A., & Sidney, S. (1994). The risk of hospitalization
for ischemic heart disease among Asian Americans in northern California. American Journal
of Public Health, 84 (10), 1672–1675.
109. Esterbauer, H., Gebicki, J., Puhl, H., & Jürgens, G. (1992). The role of lipid peroxidation and
antioxidants in oxidative modifi cation of LDL. Free Radical Biology and Medicine, 13 (4),
341–390.
110. Day, A. P., Kemp, H. J., Bolton, C., Hartog, M., & Stansbie, D. (1997). Effect of concentrated
red grape juice consumption on serum antioxidant capacity and low-density lipoprotein oxi-
dation. Annals of Nutrition and Metabolism, 41 (6), 353–357.
111. Matsumoto, H., Nakamura, Y., Hirayama, M., Yoshiki, Y., & Okubo, K. (2002). Antioxidant
activity of black currant anthocyanin aglycons and their glycosides measured by chemilumi-
nescence in a neutral pH region and in human plasma. Journal of Agricultural and Food
Chemistry, 50 (18), 5034–5037.
112. Abuja, P. M., Murkovic, M., & Pfannhauser, W. (1998). Antioxidant and prooxidant activities
of elderberry (Sambucus nigra) extract in low-density lipoprotein oxidation. Journal of
Agricultural and Food Chemistry, 46 (10), 4091–4096.
113. Rechner, A. R., & Kroner, C. (2005). Anthocyanins and colonic metabolites of dietary poly-
phenols inhibit platelet function. Thrombosis Research, 116 (4), 327–334.
114. Wang, D., Zou, T., Yang, Y., Yan, X., & Ling, W. (2011). Cyanidin-3-O-β-glucoside with the
aid of its metabolite protocatechuic acid, reduces monocyte infi ltration in apolipoprotein
E-defi cient mice. Biochemical Pharmacology, 82 (7), 713–719.
115. Hubert, P. A., Lee, S. G., Lee, S. K., & Chun, O. K. (2014). Dietary polyphenols, berries, and
age-related bone loss: A review based on human, animal, and cell studies. Antioxidants, 3 (1),
144–158.
116. Kaume, L., Gilbert, W., Smith, B. J., & Devareddy, L. (2015). Cyanidin 3-O-β-d-glucoside
improves bone indices. Journal of Medicinal Food, 18 (6), 690–697.
117. New, S. A., Robins, S. P., Campbell, M. K., Martin, J. C., Garton, M. J., Bolton-Smith, C., et
al. (2000). Dietary infl uences on bone mass and bone metabolism: Further evidence of a posi-
tive link between fruit and vegetable consumption and bone health? American Journal of
Clinical Nutrition, 71 (1), 142–151.
118. Welch, A., MacGregor, A., Jennings, A., Fairweather-Tait, S., Spector, T., & Cassidy, A.
(2012). Habitual fl avonoid intakes are positively associated with bone mineral density in
women. Journal of Bone and Mineral Research, 27 (9), 1872–1878.
119. Langsetmo, L., Hanley, D. A., Prior, J. C., Barr, S. I., Anastassiades, T., Towheed, T., et al.
(2011). Dietary patterns and incident low-trauma fractures in postmenopausal women and
men aged ≥ 50 y: A population-based cohort study. American Journal of Clinical Nutrition,
93 (1), 192–199.
120. Tanabe, S., Santos, J., La, V. D., Howell, A. B., & Grenier, D. (2011). A-type cranberry pro-
anthocyanidins inhibit the RANKL-dependent differentiation and function of human osteo-
clasts. Molecules, 16 (3), 2365–2374.
7 The Role of Anthocyanins in Health as Antioxidant, in Bone Health and as Heart…
107
121. Bickford, P. C., Tan, J., Shytle, R. D., Sanberg, C. D., El-Badri, N., & Sanberg, P. R. (2006).
Nutraceuticals synergistically promote proliferation of human stem cells. Stem Cells and
Development, 15 (1), 118–123.
122. Devareddy, L., Hooshmand, S., Collins, J. K., Lucas, E. A., Chai, S. C., & Arjmandi, B. H.
(2008). Blueberry prevents bone loss in ovariectomized rat model of postmenopausal osteo-
porosis. Journal of Nutritional Biochemistry, 19 (10), 694–699.
123. Chen, J. R., Lazarenko, O. P., Wu, X., Kang, J., Blackburn, M. L., Shankar, K., et al. (2010).
Dietary‐induced serum phenolic acids promote bone growth via p38 MAPK/β‐catenin
canonical Wnt signaling. Journal of Bone and Mineral Research, 25 (11), 2399–2411.
124. Zhang, J., Lazarenko, O. P., Blackburn, M. L., Shankar, K., Badger, T. M., & Ronis, M. J.
(2011). Feeding blueberry diets in early life prevent senescence of osteoblasts and bone loss
in ovariectomized adult female rats. PLoS One, 6 (9), e24486.
125. Dou, C., Li, J., Kang, F., Cao, Z., Yang, X., Jiang, H., et al. (2014). Dual effect of cyanidin on
RANKL‐induced differentiation and fusion of osteoclasts. Journal of Cellular Physiology .
doi:
10.1002/jcp.24916 .
126. Moriwaki, S., Suzuki, K., Muramatsu, M., Nomura, A., Inoue, F., Into, T., et al. (2014).
Delphinidin, one of the major anthocyanidins, prevents bone loss through the inhibition of
excessive osteoclastogenesis in osteoporosis model mice. PLoS One, 9 (5), e97177.
127. Watson, R., & Schönlau, F. (2015). Nutraceutical and antioxidant effects of a delphinidin-rich
maqui berry extract Delphinol
®
: A review. Minerva Cardioangiologica, 63 (2 Suppl 1), 1–12.
References
109
© The Author(s) 2016
M. Riaz et al., Anthocyanins and Human Health, SpringerBriefs in Food,
Health, and Nutrition, DOI 10.1007/978-3-319-26456-1_8
Chapter 8
The Role of Anthocyanins in Obesity
and Diabetes
8.1 Introduction
Food and phytomedicines are now becoming a trusted remedy and therapy for vari-
ous types of chronic diseases. The observed interfaces among substances present in
a phytochemical mixture strongly support the dogma of ‘eating the whole food’
rather than purifi ed compounds, extracts, nutraceuticals, or formulation. In case of
formulation, phytochemicals can loose interactions during the product development
phase leading to decreased potency of extract. As noted in previous chapters, antho-
cyanins possess a wide spectrum of biological and pharmacological effects, such as
antioxidants, anti-cancer, anti-infl ammatory, liver support by induction of detoxifi -
cation enzymes, and induction of apoptosis. As known for other phytochemicals,
mixtures of interacting anthocyanins offer boosted synergistic curative and thera-
peutic potential by multi-pronged pathways of intercession concurrently. Numerous
mechanisms of action of anthocyanins at cellular and biochemical levels can be
complementary and overlapping, and a blend of these pathways leads to the
observed health benefi ts of ingested anthocyanins. A lot of available studies indicate
potentiating interactions between various anthocyanins, and between anthocyanins
and other bioactives.
As discussed in Chaps. 1 and 7 , many studies suggest anthocyanins as promising
molecules for treatment of various diseases such as cancer, diabetes, ulcer, cardio-
vascular disorders and various metabolic disorders. While observational and lab
experimental studies appear to link dietary intake of anthocyanins to improvement
in health outcomes, data from randomized controlled trials is limited. In this regard,
two factors should be kept in mind, fi rst only small portion of ingested anthocyanins
are absorbed to exert any biological effect and second stability of anthocyanins is
regulated by various factors. Since most studies describing benefi cial effects of
anthocyanins are performed in vitro, more studies should be conducted to clarify the
mechanism of action as well as dose required to get that effect.
110
8.2 Anthocyanins and Obesity
Obesity is defi ned as an excessive accumulation of adipose tissue due to disparity
of energy intake and its disbursement. This condition is linked with several meta-
bolic disorders representing a strong risk factor for hypertension, heart disease,
hyperlipidemia and type 2 diabetes. Lack of physical activity and imbalanced
diet is a major road to obesity. Studies suggest that consumption of anthocyanins
improve the job of adipocytes and inhibit the obesity. They increase the levels of
adiponectin, an important adipocytokines, that is found at decreased levels in
obesity [ 1 ].
Animal experiments on mice confi rmed the obesity preventive effect of corns
containing anthocyanins [ 2 ]. Similar anti-obesity effects were observed in rats fed
with anthocyanins from black soybean and blueberry. Results obtained by these
experiments indicate an increase in serum high-density lipoprotein (HDL)-and a
decrease in cholesterol triglyceride and cholesterol levels [ 3 ].
Tsuda et al. (2005) studied the effect of anthocyanins on gene expression of
cultured adipocyte, it has been reported that whole blueberry results increase in
obesity while purifi ed blueberry anthocyanins has signifi cant anti-obesity action
[ 4 ]. The total RNA isolated from the adipocytes was analyzed using GeneChip
microarray. A total of 633 or 427 genes were up-regulated, after the treatment of
adipocytes with 100 μM of cyanidin-3-glucoside or cyanidin, respectively. Based
on the gene expression profi le, the up-regulation of hormone-sensitive lipase and
enhancement of the lipolytic activity were suggested to be the result of anthocy-
anin treatment on adipocytes [ 4 ]. An excessive adipose tissue (AT) accumulation
has metabolic consequences, as adipocyte dysfunction, strongly associated with
the development of obesity and diabetes, involving insulin resistance. However,
few studies suggest that AT is an important site of Vaccinium species actions to
ameliorate obesity complications [ 5 ]. Anthocyanins were shown to regulate obe-
sity and insulin sensitivity associated with adipocytokine secretion and PPARc
activation in adipocytes [ 6 ]. High fat diet along with blueberry attenuates insulin
resistance and hyperglycemia in mice coincident with reductions in adipocyte
death [ 7 ]. In another animal study, Prior et al. (2010) concluded that consumption
of purifi ed anthocyanins (0.2 mg/ml) in the drinking water (0.49 mg/mouse/day)
improved β-cell function and, the rate of fat deposition was decreased, however,
blueberry juice was not effective in preventing obesity. Interestingly enough,
lower serum leptin concentrations were found in anthocyanins treatments, which
retarded the development of obesity [
3 ].
According to Titta et al. (2009) [
8 ], orange juice has anti-obesity effect on fat
accumulation but only anthocyanins are not responsible, multiple components pres-
ent in the orange juice might act synergistically to inhibit fat accumulation. Animal
studies using Vaccinium asheii cultivars, results in reduction in food intake and,
consequently a decrease in body weight gain [ 9 ].
8 The Role of Anthocyanins in Obesity and Diabetes
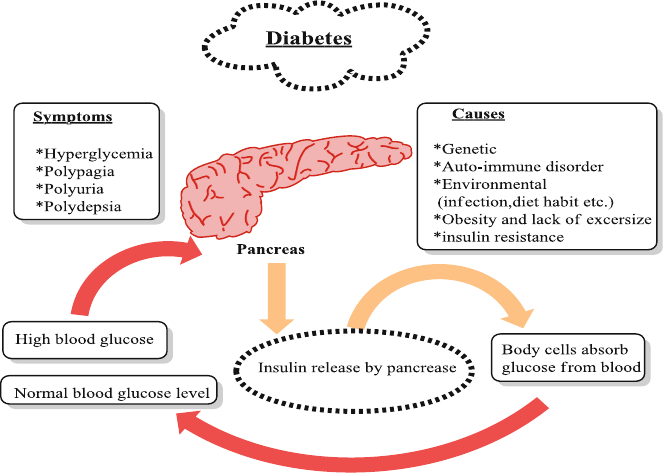
111
8.3 Anthocyanins and Diabetes
Diabetes is a metabolic disease that occurs by a combination of several genetic and
life-style patterns, which leads to hyperglycemia and associated symptoms includ-
ing polydipsia, polyuria and polyphagia (Fig. 8.1 ). Ultimately, hyperglycemia seri-
ously damages nerves and blood vessels and can lead to irritability and blurred
vision. The WHO estimates that number of diabetic patients will be doubled within
coming two decades. This exponential rise is mainly due to lack of healthy balanced
diet, sedentary life style and increasing prevalence of obesity [ 10 – 14 ].
Diabetes may arise due to persistent stress on pancreas, degeneration of β-cells,
decreased insulin secretion, and increased insulin resistance [ 15 – 19 ]. Various scien-
tists have demonstrated anti-diabetic action of anthocyanins, which can be ascribed
to several and concurrent effects of anthocyanins, including decreasing glucosuria,
Hb A1c and blood glucose, increasing insulin secretion, improving insulin resis-
tance and preventing excessive generation of free radicals [ 20 , 21 ] however plasma
insulin or blood glucose concentrations remain the same in healthy humans [ 22 ]. In
addition, anthocyanins may contribute to the prevention of type-2 diabetes through
its antioxidant activity which may protect β-cells from glucose induced oxidative
stress [ 23 ]. Sugimoto et al. (2003) reported that boysenberry anthocyanins rein-
stated or favored to reinstate the biomarkers of oxidative stress to the level of the
control group in STZ-induced diabetic rats [ 24 ].
Fig. 8.1 Pathogenesis of diabetes and glucose regulation by pancreas
8.3 Anthocyanins and Diabetes
112
Stull et al. (2010) suggested that consumption of blueberries increase insulin
action or sensitivity effectiveness in increasing insulin action in vivo, in obese and
insulin resistant (non-diabetic) human subjects, without changes in adiposity or
infl ammatory biomarkers [ 25 ]. However, more cellular mechanistic studies are
needed to clarify the specifi c cellular pathway involved in improving sensitivity to
insulin when blueberries were consumed. Furthermore, hypoglycemic activity has
been reported by various scientists as anthocyanins and anthocyanidins induce insu-
lin secretion and the order of affect observed was delphinidin-3-glucoside > cyanidin-
3- glucoside > and pelargonidin-3-galactoside [ 20 ]. Grace et al. (2009) reported
anthocyanins enriched fraction from low bush black berries for hypoglycemic activ-
ity in acute mouse model of type 2 diabetes however the fruit extract has no hypo-
glycemic effect [ 20 ]. Among these studies, Tsuda et al. (2003) evaluated that
supplementation with cyaniding-3-O-α- D -glucoside rich purple corn color improves
hyperglycemia induced by high fat diet in mice [ 26 ]. Another study by Vuong et al.
(2010) showed that biotransformed blueberry juice decreases hyperglycemia in a
diabetic animal model, by partly reversing adiponectin levels thus protects young
pre-diabetic mice from developing diabetes and obesity [ 27 ].
Tsuda et al. (2003) demonstrated that dietary anthocyanins signifi cantly normal-
ized hypertrophy of the adipocytes in the epididymal white adipose tissue with
increase in adiponectin and leptin secretion, thus diminishing hyperglycemia
infl icted by the high fat diet in mice (C57BL/6J) [ 26 ]. Later in 2005 Tsuda et al.
reported that if we treat adipocytes with cyaniding-3-glucoside or cyanidin, the hor-
mone sensitive lipase gene expression elevation occur that results in lipolytic activ-
ity and insulin sensitivity [ 4 , 26 ].
Suzuki et al. (2011) determined that bilberry extracts are capable of inhibiting
adipocyte differentiation of 3T3-L1 cells in a dose-dependent manner and to dimin-
ish lipid accumulation with concomitant down-regulation of peroxisome proliferator-
activated receptor gamma (PPARc). Additionally, all tested single anthocyanins
(pelargonidin, cyanidin, delphinidin, peonidin, and malvinidin) also inhibit lipid
accumulation in 3T3-L1 cells but delphinidin was most effective in down-regulating
PPARc and sterol regulatory element-binding protein 1c (SREBP1c) mRNA levels
[ 5 ]. Bilberry extracts inhibit adipocyte differentiation via insulin pathway and these
effects are mainly due to the presence of anthocyanins. Clinical trials involving
blueberry consumption or blueberry extract are recommended to get benefi ts of
obesity prevention and to understand it mechanism [ 9 ]. A simplifi ed way of antho-
cyanins to control diabetes is shown in Fig. 8.2 .
8.3.1 Oxidative Stress
There is a vital linkage among oxidative stress, infl ammatory response and insulin
activity. Excessive generation of free radicals occurs in hyperglycemic and hyperlipid-
emic state. In order to reduce free radicals and ultimately stress, antioxidants like
anthocyanins are required to be consumed [ 24 , 28 ]. Ischemia-reperfusion-induced
8 The Role of Anthocyanins in Obesity and Diabetes
113
oxidative stress leads to high concentrations of thiobarbituric acid reactive substances
(TBARS) and diminished levels of reduced glutathione [ 26 ], the details of anti-diabetic
studies in animals which involves reduction of oxidative stress are given in Table 8.1 .
Anthocyanins enriched freeze-dried blueberry powder increased plasma antioxi-
dant levels after ingestion of fat diet in healthy men [ 33 ]. Similarly juice prepared
from freeze-dried strawberry powder lowers lipid peroxidation, leading to decreased
plasma levels of oxidized LDL, MDA and 4-hydroxy-noneal in women with meta-
bolic syndrome after consuming two glasses for 4 weeks [ 22 ].
8.3.2 β Cells
When diabetes type 2 develops pancreatic β cells stop working due to high oxidative
stress. But anthocyanins can decrease oxidative stress by their antioxidant action
and β cells may be protected [ 31 ]. Blueberry ( Vaccinium angustifolium ) fruit extract
augmented the proliferation of pancreatic β TC-tet cells exerting a substantial anti-
diabetic effect [ 34 ] that may be due to anthocyanins [ 35 ]. Jayaprakasam et al. (2006)
Mechanism to control diabetes
β−cells
Oxidative stress
Insulin resistance
α-Glucosidase inhibitory
action
Pancreatic β TC-tet cells
Anthocyanins
pancreatic islets protection
modulation of pro-apoptotic
proteins
insulin secretion
reduced blood glucose,
TNF-α and MCP-1
inhibition of maltase
inhibition of sucrase
competitive inhibition
of
α-Glucosidase
change in the molecular
configuration
Increased GLUT4
mRNA expression
reduction in plasma
levels of MCP-1
Fig. 8.2 Mechanism to control diabetes via anthocyanins
8.3 Anthocyanins and Diabetes
114
reported that cornelian cherry anthocyanins protected the pancreatic islets of
C57BL/6 CB mice and insulin secretion was increased [ 36 ]. Treatment with antho-
cyanins from black soybean seed coats prevented the apoptosis of β cells through
modulation of pro-apoptotic proteins caspase 3 and Bax which amplifi ed Bcl2
expression which indicates anti-apoptotic effects [ 32 ]. Purifi ed anthocyanins were
reported to increase β cell function in C57BL/6J mice [ 3 ]. Anthocyanins from
Chinese bayberry extract protected pancreatic β INS-1 cells and primary islets from
damage induced by H
2
O
2
[ 37 ].
8.3.3 Insulin Secretion
In type 2 diabetes, insulin secretion may be impaired due to the dysfunction of β cells
[ 38 ]. Synthetic agents like sulfonyl urea increase insulin secretion but anthocyanins,
an important natural bioactives may act as insulin secretagogue without producing
such side effect [ 31 , 32 ]. Delphinidin-3- glucoside in INS-1 832/13 cells led to
Table 8.1 Anthocyanins that results reduction in oxidative stress
Anthocyanins Model Mechanism References
Diet containing 0.2 %
cyanidin-3-glucoside
Ischemia reperfusion
rat model
↓ Serum thiobarbituric acid
reactive substances (TBARS)
[
26 ]
↑ Hepatic reduced glutathione
in rats
Diet containing 0.1 %
mulberry anthocyanins
Streptozotocin(STZ)-
induced diabetic
Wistar rats
↓ Liver oxidized glutathione
and tended ↓ TBARS levels
[
24 ]
Diet with 0.5 % of black
rice anthocyanins-rich
extract
Sprague–Dawley rats ↓ Blood concentrations of
TBARS ↓ glutathione
[
29 ]
Diet supplemented with
0.2 % chokeberry fruit
extract
Pre-diabetic Wistar
rats
Normalize the
concentrations of TBARS in
the liver, kidneys and lungs
and no effect on SOD
[
30 ]
One i.p. injection of
pelargonidin at a dose of
3 mg/kg body weight
STZ-induced diabetic
Wistar rats
Restoration of exhausted
serum levels of SOD and
catalase and normalization
of the serum levels of MDA
[
31 ]
Anthocyanins from black
soybean seed coats
STZ-diabetic rats Reinstatement of the
depleted serum levels of
catalyze and SOD enzymes
and Normalization of the
serum levels of MDA
[
32 ]
Anthocyanins rich
ethanolic extract of
Vaccinium arctostaphylos
fruit, Orally
Alloxan-induced
diabetic Wistar rats
↑ The concentration of
glutathione peroxidase,
catalase and superoxide
dismutase in red blood cells
[
28 ]
8 The Role of Anthocyanins in Obesity and Diabetes
115
decreased insulin secretion more than cyaniding- 3-glucoside which is more potent
than pelargonidin-3-galactoside. Among various anthocyanins evaluated, only pelar-
gonidin showed secretagouge activity. Pelargonidin-3-glucoside and anthocyanins
extracted from black soybean increased insulin secretion in STZ-induced-diabetic
Sprague–Dawley and Wistar diabetic rats respectively [ 31 , 32 ]. Extract of V. arcto-
staphylos increases insulin expression in cardiac and pancreatic cells of alloxon-
induced-diabetic Wistar rats [ 28 ].
8.3.4 Insulin Resistance
Anthocyanins have been reported for anti-infl ammatory potential and can
decrease the expression of monocyte chemotactic protein-1 (MCP-1), tumor
necrosis factor α (TNF-α), interleukin-6 (IL-6), thus improving T2D and insulin
resistance [ 7 , 39 ]. The proposed hypoglycemic activity of anthocyanins can be
ascribed partly by the activation of insulin receptors generated [ 32 ]. Anthocyanins
can modify the expression of adipocytokines, intensify the expression of GLUT4,
reduce the expression of RBP4, stimulate AMPK and decrease the oxidative
stress to improve insulin sensitivity, thus positively modulating the glycemic
control. Table 8.2 indicates improvement of insulin resistance in anthocyanins
treated animal models.
8.3.5 α-Glucosidase Inhibitory Action
α-Glucosidases cause hydrolysis of carbohydrates in small intestine, thus inhibit-
ing these enzymes is one therapeutic approach to control diabetes specially after
meal [ 42 ]. Matsui et al. (2002) reported that anthocyanins extract of purple sweet
potato reduces the blood glucose through inhibition of maltase activity in healthy
Sprague–Dawley rats after carbohydrate loading (sucrose, maltose and glucose)
[ 43 ]. Another study by Jurgoński et al. (2008) reported that anthocyanins from
chokeberry lowered the high glucose level by inhibiting maltase and sucrase in
Wistar rat model of prediabetes obtained through a high-fructose diet and simulta-
neous single injection of streptozotocin (20 mg/kg) [ 30 ]. Anthocyanins cause inhi-
bition of α-glucosidases [ 43 ] but the exact mechanism of inhibition is unknown,
however it is considered to be competitive, since normal substrates of the enzyme
and glucosyl groups linked to anthocyanins are structurally similar [ 44 ]. Another
possible mechanism may be change in the molecular conformation of the enzyme
that polar groups of enzyme active site interact with hydroxyl groups on anthocya-
nins, leading to a modifi cation in enzyme activity [ 45 ]. Another important aspect to
be considered is that the use of synthetic inhibitors of α-glucosidases and anthocya-
nins might have synergistic effect and the α-glucosidases inhibition of anthocya-
nins is directly related to its stability [ 46 ].
8.3 Anthocyanins and Diabetes

116
8.3.6 Obesity and Diabetes
Due to sedentary life-style and changed dietary patterns especially large intake of
refi ned food has led to a dramatic rise in obesity and diabetes. Obesity and diabetes
are inter connected disorders and anthocyanins have been reported for health ben-
efi cial effect in association with both for example Tsuda et al. (2003, 2008) reported
high fat diet if supplemented with cyanidin-3-glucoside cause reduction in fat
Table 8.2 Insulin resistance improvement in various models
Anthocyanins Model
Insulin resistance improvement
mechanism Reference
High-fat and
anthocyanins
supplemented diet
C57BL/6J mice Reduced blood glucose, TNF-α [
26 ]
Anthocyanins
supplemented diet
KK-Ay mice Reduced blood glucose, TNF-α
and MCP-1
[
40 ]
High-fat diet
supplemented with
tart cherry powder
Zucker rats Reduced glycemia and insulinemia
as well as improved insulin
resistance due to a decrease in
plasma levels of IL-6 and TNF-α
[
39 ]
High-fat diet
supplemented with
freeze-dried whole
blueberry powder
C57BL/6 mice Reduced levels of fasting glucose
and TNF-α expression in adipose
tissue and improved insulin
tolerance test (ITT)
[
7 ]
Freeze-dried powders
from Different
anthocyanins-rich
whole berries in
hypo- and hyper-
lipidemic diets
C57BL/6 J mice No changes in plasma levels of
TNF-α, IL-6 and PAI-1
[
41 ]
High-fat diet and
drinking water
supplemented with
anthocyanins purifi ed
from blueberries
C57BL/6 J mice ↓ Plasma levels of MCP-1 [
41 ]
Anthocyanins extracted
from black soybean
Sprague–Dawley
rats
↑Autophosphorylation of insulin
receptors
[
32 ]
Anthocyanins from
different plant species
Animal models ↓ Glycemia and ↑expression of the
glucose transporter type 4 (GLUT4)
[
21 , 32 ,
40 ]
Extract of Vaccinium
arctostaphylos
Heartcells of
alloxan-induced
diabetic Wistar
rats
IncreasedGLUT4 mRNA
expression
[
28 ]
Billberry extract in diet DiabeticKK-Ay
mice
↓ Plasma glucose and ↑ insulin
sensitivity in activation of AMPK
in white adipose, skeletal muscle
and liver tissues
[
21 ]
8 The Role of Anthocyanins in Obesity and Diabetes
117
accumulation in C57BL/6J mice [ 2 , 26 ] and suppression of elevated serum glucose
levels induced by high-fat meals. Table 8.3 shows simultaneous effects of anthocy-
anin in diabetes and obesity.
8.4 Improvement of Eye Vision
The mounting evidence supported by clinical trials indicates anthocyanins as a
potential future drug to treat various opthalmological diseases. Bilberry anthocya-
nins increase night vision by interacting with rhodopsin [ 52 ] or phosphodiesterase
(PDE) on photo-transduction [ 53 , 54 ].
Berry anthocyanins improve vision by multiple mechanisms by (i) reducing
molecular degeneration and diabetic retinopathy (ii) increasing blood circulation in
retina capillaries (iii) improving night-vision by increased production of retinal pig-
ments and by (iv) preventing cataracts, retinitis pigmentosa and glaucoma [ 55 , 56 ].
Table 8.3 Anthocyanins simultaneous effects on obesity and diabetes
Anthocyanins Model Effect Reference
High-fat diet (45 % of energy)
with anthocyanins extracted
from blueberries
C57BL/6
mice
Inhibit weight gain and body fat
accumulation
[
47 ]
Whole blueberry powder C57BL/6
mice
Promote body fat accumulation [
47 ]
Blueberry juice high fat diet
(45 % of energy)
Mice Reduction in body weight gain and
percentage of white adipose tissue
[
3 ]
Supplementing a high-fat diet
(60 % of energy) with WBP
C57BL/6
mice
Did not ↓ body weight gain [
7 ]
Supplementing a high-fat diet
(45 % of energy) with 2 %
WBP
Zucker
fatty rats
Reduced abdominal fat mass and
increased the activity of the adipose
tissue and skeletal muscle
peroxisome proliferator-activated
receptor
[
48 ]
Supplementing a high-fat diet
(45 % of energy) with 2 %
WBP
Zucker
lean rats
WBP-induced body weight gain [
48 ]
Black raspberry anthocyanins
(as a juice or in powder form)
high-fat diet (60 % of energy)
Mice Didnot signifi cantly reduce body fat
accumulation or body weight gain
[
3 , 41 ,
49 ]
Mulberry water extracts high
concentration of anthocyanins
Decrease body weight gain [
50 ]
Tart cherry powder Zucker
fatty rats
Decrease body weight gain [
39 ]
Anthocyanins-rich berry,
chokeberry fructose rich diet
Mice Suppress the increase in epididymal
white adipose tissue and blood
glucose level
[
51 ]
8.4 Improvement of Eye Vision
118
Anecdotal evidence in Japan describes Royal Air Force pilots during World War
II who experienced enhanced vision in the dark after ingesting blueberry jam indi-
cating that anthocyanin ingestion can increase eye vision [ 57 ]. Decreasing the dark-
adaptation threshold was noted in a placebo-controlled cross-over double-blind
study involving healthy human subjects that were fed blackcurrant anthocyanin
concentrate [ 58 ]. The effect with the highest dose (50 mg per subject) had a statisti-
cally signifi cant effect (P = 0.011). Recently, very low (pmol g
−1
) concentration of
blueberry ANs in pig eyes were found after feeding for 4 weeks [ 59 ].
Another placebo-controlled double-blind study indicated that oral administration
of anthocyanin is vital for the production of visual purple, which helps in conversion
of light into electrical signals for brain. Adapto-electro-retinograms of two sets of
six subjects were performed before treatment and 1 and 3 h after administration; the
subjects consuming bilberries adapted to the light in 6.5 min compared to 9 min for
the control group [ 55 , 56 ]. In another clinical trial of 4 months, 50 patients suffering
from sensile cataract were given a mixture of bilberry extract, containing 25 %
anthocyanins (@180 mg twice/day) and vitamin E (@100 mg twice/day). This com-
bination stopped the development of cataract in 96 % of the subjects as compared to
76 % in control group [ 60 ].
Sundalius (2008) during her PhD research work investigated the ability of blue-
berry anthocyanins to inhibit the uptake of N -retinyl- N -retinylidene ethanolamine
(A
2
E) by retinal pigment epithelial (RPE) cells as quantifi ed by pigment epithelial
derived factor (PEDF) levels in RPE cells. The average PEDF results showed that
blueberry anthocyanins stimulated and sustained PEDF levels as compared to con-
trol RPE cells and RPE cells treated with A
2
E thus it was evaluated that blueberry
anthocyanins preserved RPE cells from oxidation as indicated by PEDF values [ 61 ].
Bilberry anthocyanins have been shown to exhibit neuroprotective effects in ocu-
lar tissue and suppressive effects on diabetic retinopathy via a reduction of angio-
genesis [ 62 , 63 ]. Furthermore, several studies have demonstrated that blackcurrant
anthocyanins improve visual function; for example, BA inhibit transient myopia,
reduce eye fatigue, improve dark adaptation, and increase retinal blood fl ow in glau-
coma [ 58 , 64 ]. The fi rst three benefi ts have been shown in humans, and effective
daily BA intake was reported as 50 mg [ 58 ]. Two studies have indicated the possible
mechanism underlying inhibition of myopia. The blackcurrant anthocyanins con-
centration was higher in ocular tissues (sclera, choroid, ciliary body, retina, iris, and
cornea) than in blood in a rat fed a diet containing blackcurrant anthocyanins, and
in particular, concentrations in the sclera and choroid were 100-fold higher than the
blood blackcurrant anthocyanins concentration [ 65 ]. In addition, it was shown that
an anthocyanins in the concentration range of 10
−7
to 10
−8
M relaxed endothelin- 1-
induced contraction of bovine ciliary smooth muscle [ 66 ], which plays an important
role in modulating refraction of the lens through contraction and relaxation, which
in turn control accommodation. Thus, it is suggested that anthocyanins stimulate the
endothelin-1 receptor to induce production of NO, thereby relaxing ciliary smooth
muscle, which in turn fl attens the lens and consequently inhibits myopia.
Vaccinium myrtillus extracts containing anthocyanins were reported to improve
the night vision, particular diabetic retinopathy, through the infl uence on the perme-
8 The Role of Anthocyanins in Obesity and Diabetes
119
ability of retinal vessels which also decrease the permeability of the blood-brain bar-
rier to toxic substances [ 67 ]. The differences in results and conclusions are probably
due to variations in dose and source of anthocyanin, type of subjects and the tech-
niques used to evaluate vision.
8.5 Conclusions
Health is a key priority of people of all age and income group and a vital feature of
a prosperous life. Health-concerned consumers of modern society are now moving
from curative to preventive health measures. It has led to the generation of dietary
recommendation and food composition tables. It is well- known now that plant-
based foods and bioactive compounds found in them like anthocyanins help in
remaining healthy. Anthocyanins act as preventive and curative agents for various
diseases and disorders as presented in this chapter. Works that suggested protective
actions of anthocyanins are mainly based on in vitro experiments, a fact that com-
promises the robustness of these fi ndings. More details are required to establish the
real implications of anthocyanins and the mechanism by which they exert their bio-
logical effects.
References
1. Arita, Y., Kihara, S., Ouchi, N., Takahashi, M., Maeda, K., Miyagawa, J., et al. (1999).
Paradoxical decrease of an adipose-specifi c protein, adiponectin, in obesity. Biochemical and
Biophysical Research Communications, 257 (79–83), 466.
2. Tsuda, T. (2008). Regulation of adipocyte function by anthocyanins; possibility of preventing
the metabolic syndrome. Journal of Agricultural and Food Chemistry, 56 (3), 642–646.
3. Prior, R. L., Wilkes, S. E., Rogers, T. R., Khanal, R. C., Wu, X., & Howard, L. R. (2010).
Purifi ed blueberry anthocyanins and blueberry juice alter development of obesity in mice fed
an obesogenic high-fat diet. Journal of Agricultural and Food Chemistry, 58 (7), 3970–3976.
4. Tsuda, T., Ueno, Y., Kojo, H., Yoshikawa, T., & Osawa, T. (2005). Gene expression profi le of
isolated rat adipocytes treated with anthocyanins. Biochimica et Biophysica Acta, 1733 (2),
137–147.
5. Suzuki, R., Tanaka, M., Takanashi, M., Hussain, A., Yuan, B., Toyoda, H., et al. (2011).
Anthocyanidins-enriched bilberry extracts inhibit 3T3-L1 adipocyte differentiation via the
insulin pathway. Nutrition and Metabolism, 8 (1), 14.
6. Tsuda, M., Mizokoshi, A., Shigemoto-Mogami, Y., Koizumi, S., & Inoue, K. (2004). Activation
of p38 mitogen activated protein kinase in spinal hyperactive microglia contributes to pain
hypersensitivity following peripheral nerve injury. Glia, 45 (1), 89–95.
7. DeFuria, J., Bennett, G., Strissel, K. J., Perfi eld, J. W., Milbury, P. E., Greenberg, A. S., et al.
(2009). Dietary blueberry attenuates whole-body insulin resistance in high fat-fed mice by
reducing adipocyte death and its infl ammatory sequelae. Journal of Nutrition, 139 (8),
1510–1516.
8. Titta, L., Trinei, M., Stendardo, M., Berniakovich, I., Petroni, K., Tonelli, C., et al. (2009).
Blood orange juice inhibits fat accumulation in mice. Int J Obesity, 34 (3), 578–588.
References
120
9. Norberto, S., Sara, S., Manuela, M., Ana, F., Manuela, P., & Conceição, C. (2013). Blueberry
anthocyanins in health promotion: A metabolic overview. Journal of Functional Foods, 5 (4),
1518–1528.
10. Saad, B., & Said, O. (2011). Greco-Arab and Islamic herbal medicine: Traditional system,
ethics, safety, effi cacy, and regulatory issues . Hoboken: Wiley.
11. Saad, B., Zaid, H., & Said, O. (2012). Tradition and perspectives of diabetes treatment in
Greco-Arab and Islamic medicine. In Bioactive food as dietary interventions for diabetes:
Bioactive foods in chronic disease states (pp. 319–326).
12. Zaid, H., & Saad, B. (2012). State of the art of diabetes treatment in Greco-Arab and Islamic
medicine. In Bioactive food as dietary interventions for diabetes: Bioactive foods in chronic
disease states (p. 327).
13. Saad, B. (2015). Integrating traditional Greco‐Arab and Islamic diet and herbal medicines in
research and clinical practice. In Phytotherapies: Effi cacy, safety, and regulation (p. 142).
14. Kadan, S., Saad, B., Sasson, Y., & Zaid, H. (2013). In vitro evaluations of cytotoxicity of eight
antidiabetic medicinal plants and their effect on GLUT4 translocation. Evidence-Based
Complementary and Alternative Medicine, 2013 .
15. Sultan, M. T., Butt, M. S., Karim, R., Iqbal, S. Z., Ahmad, S., Zia-Ul-Haq, M., et al. (2014).
Effect of Nigella sativa fi xed and essential oils on antioxidant status, hepatic enzymes, and
immunity in streptozotocin induced diabetes mellitus. BMC Complementary and Alternative
Medicine, 14 (1), 193.
16. Bukhari, S. A., Shamshari, W. A., Ur-Rahman, M., Zia-Ul-Haq, M., & Jaafar, H. Z. (2014).
Computer aided screening of secreted frizzled-related protein 4 (SFRP4): A potential control
for diabetes mellitus. Molecules, 19 (7), 10129–10136.
17. Khanra, R., Dewanjee, S., Dua, T. K., Sahu, R., Gangopadhyay, M., De Feo, V., et al. (2015).
Abroma augusta L. (Malvaceae) leaf extract attenuates diabetes induced nephropathy and car-
diomyopathy via inhibition of oxidative stress and infl ammatory response. Journal of
Translational Medicine, 13 (1), 1–14.
18. Sultan, M. T., Butt, M. S., Karim, R., Zia-Ul-Haq, M., Batool, R., Ahmad, S., et al. (2014).
Nigella sativa fi xed and essential oil supplementation modulates hyperglycemia and allied
complications in streptozotocin-induced diabetes mellitus. Evidence-Based Complementary
and Alternative Medicine, 2014 .
19. Zia-Ul-Haq, M., Ahmad, S., Bukhari, S. A., Amarowicz, R., Ercisli, S., & Jaafar, H. Z. (2014).
Compositional studies and biological activities of some mash bean ( Vigna mungo (L.) Hepper)
cultivars commonly consumed in Pakistan. Biological Research, 47 , 23.
20. Grace, M. H., Ribnicky, D. M., Kuhn, P., Poulev, A., Logendra, S., Yousef, G. G., et al. (2009).
Hypoglycemic activity of a novel anthocyanin-rich formulation from lowbush blueberry,
Vaccinium angustifolium Aiton. Phytomedicine, 16 (5), 406–415.
21. Takikawa, M., Inoue, S., Horio, F., & Tsuda, T. (2010). Dietary anthocyanin-rich bilberry
extract ameliorates hyperglycemia and insulin sensitivity via activation of AMP-activated pro-
tein kinase in diabetic mice. Journal of Nutrition, 140 (3), 527–533.
22. Basu, A., & Penugonda, K. (2009). Pomegranate juice: A heart‐healthy fruit juice. Nutrition
Reviews, 67 (1), 49–56.
23. Al-Awwadi, N. A., Araiz, C., Bornet, A., Delbosc, S., Cristol, J. P., Linck, N., et al. (2005).
Extracts enriched in different polyphenolic families normalize increased cardiac NADPH oxi-
dase expression while having differential effects on insulin resistance, hypertension, and car-
diac hypertrophy in high-fructose-fed rats. Journal of Agricultural and Food Chemistry, 53 (1),
151–157.
24. Sugimoto, M., Kuo, M. L., Roussel, M. F., & Sherr, C. J. (2003). Nucleolar Arf tumor suppres-
sor inhibits ribosomal RNA processing. Molecular Cell, 11 (2), 415–424.
25. Stull, A. J., Cash, K. C., Johnson, W. D., Champagne, C. M., & Cefalu, W. T. (2010). Bioactives
in blueberries improve insulin sensitivity in obese, insulin-resistant men and women. Journal
of Nutrition, 140 (10), 1764–1768.
26. Tsuda, T., Horio, F., Uchida, K., Aoki, H., & Osawa, T. (2003). Dietary cyanidin 3-O-β-D-
glucoside-rich purple corn color prevents obesity and ameliorates hyperglycemia in mice.
Journal of Nutrition, 133 (7), 2125–2130.
8 The Role of Anthocyanins in Obesity and Diabetes
121
27. Vuong, T., Matar, C., Ramassamy, C., & Haddad, P. S. (2010). Biotransformed blueberry juice
protects neurons from hydrogen peroxide-induced oxidative stress and mitogen-activated pro-
tein kinase pathway alterations. British Journal of Nutrition, 104 (05), 656–663.
28. Feshani, A. M., Kouhsari, S. M., & Mohammadi, S. (2011). Vaccinium arctostaphylos, a com-
mon herbal medicine in Iran: Molecular and biochemical study of its antidiabetic effects on
alloxan-diabetic Wistar rats. Journal of Ethnopharmacology, 133 (1), 67–74.
29. Guo, H., Ling, W., Wang, Q., Liu, C., Hu, Y., Xia, M., et al. (2007). Effect of anthocyanin-rich
extract from black rice ( Oryza sativa L. indica) on hyperlipidemia and insulin resistance in
fructose-fed rats. Plant Foods for Human Nutrition, 62 (1), 1–6.
30. Jurgoński, A., Juśkiewicz, J., & Zduńczyk, Z. (2008). Ingestion of black chokeberry fruit
extract leads to intestinal and systemic changes in a rat model of prediabetes and hyperlipid-
emia. Plant Foods for Human Nutrition, 63 (4), 176–182.
31. Roy, M., Sen, S., & Chakraborti, A. S. (2008). Action of pelargonidin on hyperglycemia and
oxidative damage in diabetic rats: Implication for glycation-induced hemoglobin modifi cation.
Life Sciences, 82 (21), 1102–1110.
32. Nizamutdinova, I. T., Kim, Y. M., Chung, J. I., Shin, S. C., Jeong, Y. K., Seo, H. G., et al.
(2009). Anthocyanins from black soybean seed coats stimulate wound healing in fi broblasts
and keratinocytes and prevent infl ammation in endothelial cells. Food and Chemical Toxicology,
47 (11), 2806–2812.
33. Kay, C. D., & Holub, B. J. (2002). The effect of wild blueberry ( Vaccinium angustifolium )
consumption on postprandial serum antioxidant status in human subjects. British Journal of
Nutrition, 88 (04), 389–397.
34. Martineau, L. C., Couture, A., Spoor, D., Benhaddou-Andaloussi, A., Harris, C., Meddah, B.,
et al. (2006). Anti-diabetic properties of the Canadian lowbush blueberry Vaccinium angusti-
folium Ait. Phytomedicine, 13 (9), 612–623.
35. Prior, R. L., Cao, G., Martin, A., Sofi c, E., McEwen, J., O’Brien, C., et al. (1998). Antioxidant
capacity as infl uenced by total phenolic and anthocyanin content, maturity, and variety of
Vaccinium species. Journal of Agricultural and Food Chemistry, 46 (7), 2686–2693.
36. Jayaprakasam, B., Olson, L. K., Schutzki, R. E., Tai, M. H., & Nair, M. G. (2006). Amelioration
of obesity and glucose intolerance in high-fat-fed C57BL/6 mice by anthocyanins and ursolic
acid in Cornelian cherry ( Cornus mas ). Journal of Agricultural and Food Chemistry, 54 (1),
243–248.
37. Zhang, B., Kang, M., Xie, Q., Xu, B., Sun, C., Chen, K., et al. (2010). Anthocyanins from
Chinese bayberry extract protect β cells from oxidative stress-mediated injury via HO-1 upreg-
ulation. Journal of Agricultural and Food Chemistry, 59 (2), 537–545.
38. Ahrén, B., Pacini, G., Foley, J. E., & Schweizer, A. (2005). Improved meal-related β-cell func-
tion and insulin sensitivity by the dipeptidyl peptidase-IV inhibitor vildagliptin in metformin-
treated patients with type 2 diabetes over 1year. Diabetes Care, 28 (8), 1936–1940.
39. Seymour, E. M., Lewis, S. K., Urcuyo-Llanes, D. E., Tanone, I. I., Kirakosyan, A., Kaufman,
P. B., et al. (2009). Regular tart cherry intake alters abdominal adiposity, adipose gene tran-
scription, and infl ammation in obesity-prone rats fed a high fat diet. Journal of Medicinal
Food, 12 (5), 935–942.
40. Sasaki, R., Nishimura, N., Hoshino, H., Isa, Y., Kadowaki, M., Ichi, T., et al. (2007). Cyanidin
3-glucoside ameliorates hyperglycemia and insulin sensitivity due to down regulation of reti-
nol binding protein 4 expression in diabetic mice. Biochemical Pharmacology, 74 (11),
1619–1627.
41. Prior, R. L., Wu, X., Gu, L., Hager, T., Hager, A., Wilkes, S., et al. (2009). Purifi ed berry
anthocyanins but not whole berries normalize lipid parameters in mice fed an obesogenic high
fat diet. Molecular Nutrition and Food Research, 53 (11), 1406–1418.
42. Chiasson, J. L., Josse, R. G., Gomis, R., Hanefeld, M., Karasik, A., Laakso, M., et al. (2002).
Acarbose for prevention of type 2 diabetes mellitus: The STOP-NIDDM randomised trial. The
Lancet, 359 (9323), 2072–2077.
43. Matsui, T., Ebuchi, S., Kobayashi, M., Fukui, K., Sugita, K., Terahara, N., et al. (2002). Anti-
hyperglycemic effect of diacylated anthocyanin derived from Ipomoea batatas cultivar
References
122
Ayamurasaki can be achieved through the α-glucosidase inhibitory action. Journal of
Agricultural and Food Chemistry, 50 (25), 7244–7248.
44. McDougall, G. J., & Stewart, D. (2005). The inhibitory effects of berry polyphenols on diges-
tive enzymes. Biofactors, 23 (4), 189–195.
45. Adisakwattana, S., Charoenlertkul, P., & Yibchok-anun, S. (2009). α-Glucosidase inhibitory
activity of cyanidin-3-galactoside and synergistic effect with acarbose. Journal of Enzyme
Inhibition and Medicinal Chemistry, 24 (1), 65–69.
46. Sancho, R. A. S., & Pastore, G. M. (2012). Evaluation of the effects of anthocyanins in type 2
diabetes. Food Research International, 46 (1), 378–386.
47. Prior, R. L., Wu, X., Gu, L., Hager, T. J., Hager, A., & Howard, L. R. (2008). Whole berries
versus berry anthocyanins: Interactions with dietary fat levels in the C57BL/6 J mouse model
of obesity. Journal of Agricultural and Food Chemistry, 56 (3), 647–653.
48. Seymour, E. M., Tanone, I. I., Urcuyo-Llanes, D. E., Lewis, S. K., Kirakosyan, A., Kondoleon,
M. G., et al. (2011). Blueberry intake alters skeletal muscle and adipose tissue peroxisome
proliferator-activated receptor activity and reduces insulin resistance in obese rats. Journal of
Medicinal Food, 14 (12), 1511–1518.
49. Kaume, L., William, C. G., Cindi, B., Luke, R. H., & Latha, D. (2012). Cyanidin 3-O-β-D-
glucoside-rich blackberries modulate hepatic gene expression, and anti-obesity effects in
ovariectomized rats. Journal of Functional Foods, 4 (2), 480–488.
50. Peng, C.-H., Liu, L. K., Chuang, C. M., Chyau, C. C., Huang, C. N., & Wang, C. J. (2011).
Mulberry water extracts possess an anti-obesity effect and ability to inhibit hepatic lipogenesis
and promote lipolysis. Journal of Agricultural and Food Chemistry, 59 (6), 2663–2671.
51. Qin, B., & Anderson, R. A. (2012). An extract of chokeberry attenuates weight gain and modu-
lates insulin, adipogenic and infl ammatory signalling pathways in epididymal adipose tissue of
rats fed a fructose-rich diet. British Journal of Nutrition, 108 (04), 581–587.
52. Bastide, P., Rouher, F., & Tronche, P. (1967). Rhodopsin and anthocyanosides. Apropos of
various experimental facts. Bulletin des Sociétés d’Ophtalmologie de France, 68 (9),
801–807.
53. Ferretti, C., Magistretti, M. J., Robotti, A. P., & Genazzani, G. E. (1988). Vaccinium myrtillus
anthocyanosides are inhibitors of cAMP and cGMP phosphodiesterases. Pharmacological
Research Communications, 20 , 150.
54. Virmaux, N., Bizec, J. C., Nullans, G., Ehret, S., & Mandel, P. (1990). Modulation of rod
cyclic GMP-phosphodiesterase activity by anthocyanidin derivatives. Biochemical Society
Transactions, 18 (4), 686–687.
55. Camire, M. E. (2000). Bilberries and blueberries as functional foods and nutraceuticals. In G.
Mazza & B. D. Oomah (Eds.), Functional foods: Herbs, botanicals and teas (pp. 289–319).
Lancaster: Technomic.
56. Camire, M. (2000). In G. Mazza & B. D. Oomah (Eds.), Functional foods: Herbs, botanicals
and teas . Lancaster: Technomic.
57. Canter, P. H., & Ernst, E. (2004). Anthocyanosides of Vaccinium myrtillus (Bilberry) for night
vision-a systematic review of placebo-controlled trials. Survey of Ophthalmology, 49 (1),
38–50.
58. Nakaishi, H., Matsumoto, H., Tominaga, S., & Hirayama, M. (2000). Effects of black currant
anthocyanoside intake on dark adaptation and VDT work-induced transient refractive altera-
tion in healthy humans. Alternative Medicine Review, 5 (6), 553–562.
59. Kalt, W., Blumberg, J. B., McDonald, J. E., Vinqvist-Tymchuk, M. R., Fillmore, S. A., Graf,
B. A., et al. (2008). Identifi cation of anthocyanins in the liver, eye, and brain of blueberry-fed
pigs. Journal of Agricultural and Food Chemistry, 56 (3), 705–712.
60. Head, K. (2001). Natural therapies for ocular disorders part two: Cataracts and glaucoma.
Alternative Medicine Review, 6 (2), 141–166.
61. Sundalius, N. M. (2008). Examination of blueberry anthocyanins in prevention of age-related
macular degeneration through retinal pigment epithelial cell culture study . Faculty of the
Louisiana State University and Agricultural and Mechanical College in partial fulfi llment of
8 The Role of Anthocyanins in Obesity and Diabetes
123
the requirements for the degree of Master of Science in The Department of Food Science by
Naomi Marie Sundalius BS, Michigan State University.
62. Matsunaga, N., Tsuruma, K., Shimazawa, M., Yokota, S., & Hara, H. (2010). Inhibitory
actions of bilberry anthocyanidins on angiogenesis. Phytotherapy Research, 24 (S1),
S42–S47.
63. Matsunaga, N., Chikaraishi, Y., Shimazawa, M., Yokota, S., & Hara, H. (2010). Vaccinium
myrtillus (bilberry) extracts reduce angiogenesis in vitro and in vivo. Evidence-Based
Complementary and Alternative Medicine, 7 (1), 47–56.
64. Iida, H., Nakamura, Y., Matsumoto, H., Takeuchi, Y., Harano, S., Ishihara, M., et al. (2010).
Effect of black-currant extract on negative lens-induced ocular growth in chicks. Ophthalmic
Research, 44 (4), 242–250.
65. Matsumoto, H., Nakamura, Y., Iida, H., Ito, K., & Ohguro, H. (2006). Comparative assessment
of distribution of blackcurrant anthocyanins in rabbit and rat ocular tissues. Experimental Eye
Research, 83 (2), 348–356.
66. Matsumoto, H., Kamm, K. E., Stull, J. T., & Azuma, H. (2005). Delphinidin-3-rutinoside
relaxes the bovine ciliary smooth muscle through activation of ET B receptor and NO/cGMP
pathway. Experimental Eye Research, 80 (3), 313–322.
67. Igarashi, Y., Chiba, H., Utsumi, H., Miyajima, H., Ishizaki, T., Gotoh, T., et al. (2000).
Expression of receptors for glial cell line-derived neurotrophic factor (GDNF) and neurturin in
the inner blood-retinal barrier of rats. Cell Structure and Function, 25 (4), 237–241.
References
125
© The Author(s) 2016
M. Riaz et al., Anthocyanins and Human Health, SpringerBriefs in Food,
Health, and Nutrition, DOI 10.1007/978-3-319-26456-1_9
Chapter 9
Anthocyanins Effects on Carcinogenesis,
Immune System and the Central Nervous
System
9.1 Introduction
As mentioned in Chap. 1 , anthocyanins are phytochemicals that are not required for
the immediate survival of the plant but which are synthesized to increase the fi tness
of the plants to survive by allowing them to interact with their environment, includ-
ing pathogens and herbivorous and symbiotic insects. In many cases, the effects of
these secondary metabolites on the human immune system and central nervous sys-
tem might be linked either to their ecological roles in the life of the plants or to
molecular and biochemical similarities in the biology of plants and higher animals.
The health professionals now recognize that in addition to the macromolecules, i.e.
carbohydrates, lipids and proteins, there is dearth of phytochemicals like anthocya-
nins and carotenoids which have major heath promoting effects. Age-related disor-
ders like hypertension, diabetes, cardiovascular diseases, Alzheimer’s disease,
cataracts, neurodegenerative problems and macular degeneration, and improvement
of vision and brain functions may be prevented by taking ample amount of these
bioactive constituents.
9.2 Anti-Infl ammatory Activity
Infl ammation is a multifaceted biological response of vascular tissue to stimulants, irri-
tants or injuries and is linked with instigation and progression of various chronic dis-
eases like cardiovascular diseases, Alzheimer’s, disease, diabetes mellitus (type 2) and
various types of cancers [ 1 – 9 ]. Therefore, anti-infl ammatory agents also can act as
anticancer agents. Anti-infl ammatory properties of anthocyanins, anthocyadin (puri-
fi ed) and their concerned sources were reported by various scientists. For example,
Cy-aglycone was found to exhibits higher anti-infl ammatory effects than aspirin in
COX-assays [ 10 ]. Purifi ed anthocyanin fractions from bilberries, blueberries,
126
blackberries, cranberries, sweet cherries, raspberries, elderberries, strawberries and tart
cherries exhibit anti-infl ammatory properties as assessed by COX-1 and COX-2 inhibi-
tory assays and it may be due to cyanidin glycosides [ 11 ]. Strawberry, blackberry, and
raspberry showed the highest anti-infl ammatory activity, comparable to that of ibupro-
fen and naproxen at 10 μM concentrations. In an in vivo study, the therapeutic effi cacy
of blackberry anthocyanins (Cyanidin-3-glucoside accounts for 80 %) was investigated
in rats with carrageenan-induced lung infl ammation [ 12 ]. Anthocyanins effectively
reduced all parameters of infl ammation dose-dependently.
Anthocyanidins possess structure dependent anti-infl ammatory properties. The
action mechanism of this effect is believed to be mediated through inhibiting
COX-2 in lipo-poly-saccharide (LPS)-activated cells (RAW-264) or inhibiting
inducible nitric acid synthase (iNOS) and mRNA expression in LPS-activated
murine-J774 macrophages [ 13 ]. Cyclooxygenase-2 is involved in many infl amma-
tory actions. Delphinidinis are the strongest inhibitor of COX-2 expression at
mRNA and protein levels and exerts inhibitory effect on degradation of nuclear
translocation of p65 and IκB-α [ 14 ].
Anthocyanins or anthocyanin-containing extract inhibit pro-infl ammatory cyto-
kines in vitro by suppressing NF-κB through down-regulation of mitogen-activated
protein kinase (MAPK) pathways [ 15 ]. Some authors reported that these actions are
due to antioxidant effect but some authors reported anthocyanins can exhibit sub-
stantial anti-infl ammatory property through signaling pathways responsible for
anti-infl ammatory action without affecting the in vivo anti-oxidative status or its
signaling pathways [ 16 , 17 ]. Wang and his colleagues [ 17 ] found that cyanidin- 3-
glucoside inhibit COX-2 expression and iNOS by induction of liver-X-receptor-α
(LRX-α) activation in THP-1 macrophages. The signaling pathways of the nuclear
receptors are activated by anthocyanins effectively, for example LRX α and PPAR γ
(peroxisome proliferator-activated receptor γ) that antagonizes in vitro infl amma-
tory gene expression [ 14 ]. In carrageenan-induce infl ammation in rat lungs, blue-
berry anthocyanins reduced all infl ammation parameters dose-dependently [ 12 ].
Anthocyanins obtained from red wine inhibited TNF-α-induced infl ammation by
modulating the endothelial monocyte chemo-attractant protein-1 [ 18 ].
9.3 Anthocyanins and Cancer
Cancer is a principal reason of causality and mortality globally. Numerical fi gures
specify that cancer affects more than 1/3rd of world population and more than 20 %
death occur due to this malady. In both benign and malignant cancers, cells divide
abnormally by escaping the regulatory control of cell resulting in invasion of other
body tissues. Till now, more than 100 various type of cancer have been discovered
which have been classifi ed depending upon affected tissue or organ like blood can-
cer, colon cancer and breast cancer etc. Cancer results from DNA abnormalities
Infl ammation, unhealthy diet and stress-induced oxidative damages are one of major
reasons of cancer. Diet and phytochemicals have been used since dawn of civilization
9 Anthocyanins Effects on Carcinogenesis, Immune System and the Central Nervous…
127
as therapeutic strategy to cure various diseases including cancer. Bioactive constitu-
ents present in food commodities like grains, fruits, vegetables, nuts, legumes and
herbs encompass effective protection against various types of cancers. Therefore
dietary chemo-prevention is now focus of attention globally [ 19 – 21 ].
Anthocyanins, although discovered in 1918, their anticancer effects have been
studied and reported only recently. Table 9.1 lists anticancer effects of various antho-
cyanins. Modulation of carcinogenesis occurs through variety of biological func-
tions of anthocyanins. Anthocyanins are believed to exert anti-cancer effects by
multiple mechanisms like by (i) arresting cell cycle by arresting the G1/G0 and
G2/M phase (ii) inducing apoptosis and anti-angiogenesis (iii) inhibiting oxidative
DNA damage (iv) inducing phase II enzymes for detoxifi cation and (v) by inhibiting
COX-2 enzymes. The chemopreventive action of anthocyanins is due to their anti-
oxidant property.
Anthocyadins are more effective in inhibition of cell proliferation than anthocya-
nins [ 40 ]. The latter are more potent anti-cancer agents than other fl avonoids [ 23 ].
In another study by Kamei and his colleagues, anthocyanins from red wine showed
higher anticancer activity than other fl avonoids of red or white wine [ 24 ].
Anthocyanins fraction from 4 cultivars of muscadine grape exhibited higher antip-
roliferative activity than crude extract or phenolic acid fraction [ 28 ]. Antiproliferative
effects of anthocyanins on the colon cancer cells are structure dependant [ 27 ]. The
proposed mechanism of anticancer effects of anthocyanins is shown in Fig. 9.1 .
The work of Jing et al. [ 27 ] advocated the additive effect of anthocyanins and
other phenolics in combinational usage in antiproliferative studies. Various animal
studies showed that anthocyanins have chemopreventive effects in gastrointestinal
tract like oral cavity, the esophagus [ 41 ], and the colon [ 30 ]. This chemopreventive
effect may be due to anthocyanins contact directly with the epithelial layer [ 42 ]
while non-gastrointestinal organs required anthocyanins availability through blood
delivery. It was confi rmed by strawberry anthocyanins failure to inhibit
4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-and benzo[a]pyrene-induced lung
cancer in a mice model [ 43 ].
Anti-cancer activity of anthocyanins may be attributed to the additive effect of
multiple mechanisms [ 30 ]. Possible mechanisms that have been suggested are given
in Table 9.2 . Various studies showed that anthocyanins can inhibit the growth of
different cancer cells and embryonic fi broblasts, indicating their potential as chemo-
preventive agents in the form of cheap and safe anticancer dietary supplements [ 50 ].
Anthocyanins exert antiproliferative activity against human cancer cells derived
from malignant tissues [ 40 ]. Anthocyanins were also potent and selective
proliferation- inhibitor of human promyelocytic leukemia cells [ 51 ]. The inhibitory
effect on cancer cells of berry extracts deepens upon the contents and not the com-
position of anthocyanins [ 52 ].
Antiangiogenic effect, increased apoptosis and decreased proliferation were
observed in patients suffering from colon cancer administered black raspberry pow-
der daily for several weeks [ 16 ]. In another clinical study, 25 colorectal cancer
patients were given standardized anthocyanin bilberries extract (mirtocyan)
administered daily for 1 week, decrease in proliferation index, and increase in apop-
9.3 Anthocyanins and Cancer

128
Table 9.1 Anticancer activities reported from different anthocyanins
Anthocyanins source Model Effect Reference
Anthocyanin-rich
extract from
chokeberry
Colon cancer HT-29 cells Induce cell cycle
block at G1/G0 and
G2/M phases but not
in NCW460 normal
colonic cells
[
22 ]
Anthocyanins
extracted from
fl ower petals
Intestinal carcinoma derived
HCT-15 cell line
Cell growth
inhibition
[
23 ]
Anthocyanins from
red wine or white
wine
Humangastric cancer HCT-15
cell line and AGS cell line
Cell growth
inhibition
[
24 ]
Commercial bilberry,
chokeberry, and grape
extracts
HT-29 cell line Cell growth
inhibition
[
25 ]
Four anthocyanins
isolated from
strawberry
Human oral (CAL-27, KB), colon
(HT29, HCT-116), and prostate
(LNCaP, DU145) cancer cells
Reduction in cell
viability
[
26 ]
Anthocyanins rich
extract of purple corn,
purple carrot, and red
radishes
HT-29 cell line Cell growth
inhibition
[
27 ]
Anthocyanins fraction
from muscadine
grapes (4 cultivars)
Human colon cancer derived cell
liens, HT-29 and Caco-2
Cell growth
inhibition
[
28 ]
Cranberry extract
versus its fl avonol
glycosides (gly),
anthocyanins,
proanthocyanidins,
and organic acids
fractions
Human oral (KB, CAL27), colon
(HT-29, HCT116, SW480,
SW620), and prostate (RWPE-1,
RWPE-2, 22Rv1) cancer cell
lines
Anthocyanins and the
proanthocyanidin
fraction exhibited
substantial inhibitory
effect except SW480
cell lines, but
combination of both
exhibited inhibition
of all comparatively
[
29 ]
Dietary anthocyanins Colon carcinogen azoxymethane
(AOM)-induced rat colon cancer
model
Inhibitory effect on
cell proliferartion
[
30 ]
Anthocyanins rich
extracts of grapes,
bilberries and
chokeberries
Human malignant HT-29 colon
cancer cells
Cell growth
inhibition
[
25 ]
Anthocyanins rich
extracts of grapes,
bilberries and
chokeberries
Low and high tumorigenic colon
cancer cell lines, LoVo/Adr and
LoVo
Cell growth
inhibition
[
31 ]
(continued)
9 Anthocyanins Effects on Carcinogenesis, Immune System and the Central Nervous…

129
totic index was observed [ 53 ]. No substantial association was observed between
anthocyanidins intake and risk for pharyngeal or oral cancer [
54 ]. Similarly no pro-
tective effect has been reported against the development of prostate cancer [ 55 ]. To
get optimum results the anthocyanins containing berry are consumed before, during
and after chemotherapy of cancer may maximize chemopreventive effectiveness in
humans. The difference in response against various tumors may be due to its bioaval-
ibility to the site [
56 ]. Studies are required to evaluate different anthocyanins in com-
bination with various chemotherapeutic agents to maximize rational treatment of
cancer. Anthocyanins from highbush blueberry and mulberry fruits ( V. angustifolium )
stopped the proliferation activities of cells [ 57 ]. Delphinidin inhibited invasion of
human fi brosarcoma cells by down regulation of gene expression of MMP-2/9 [ 58 ].
Delphinin is more potent antioxidant due to highest number of OH groups on B ring.
Table 9.1 (continued)
Anthocyanins source Model Effect Reference
Anthocyanins from
tart cherries
Human colon cancer cells HT29
and HCT-116
Reduced proliferation [
32 ]
Anthocyanins extract
from Vaccinium
uliginosum
Human colorectal cancer cells
DLD-1 and COLO205
Growth suppression
in a dose-dependent
manner
[
33 ]
Red grape pomace
extract (oenocyanin)
Adenoma development in the
ApcMin mouse
Suppress adenoma
cell proliferation and
↓-regulation of
expression of the PI3
pathway component
Akt,
[
34 ]
Vaccinium
myrtillus , Vaccinium
vitis - idaea and Rubus
chamaemorus , rich in
anthocyanins
Min/1 mice Chemopreventive as
signifi cant ↓ in the
number of intestinal
tumors
[
35 ]
Grape juice
comprising 15
different anthocyanins
Rats mammary tumors Incidence,
multiplicity and fi nal
mass reduction
[
36 ]
Lyophilized black
raspberries
NMBA
(Nitrosomethylbenzylamine)-
induced esophageal tumors
Prevention of tumor
development
[
37 ]
Anthocyanins
containing
pomegranate extract
DMBA
(7,12-dimethylbenzanthracene)-
induced skin tumors in CD-1
mice.
Deferred the onset
and reduced the
incidence of
[
16 ]
Black rice
anthocyanins
Human colon cancer cells HT-29
and HCT-116
Reduced the
expression of MMP-2
and MMP-9 in cancer
cells
[
38 ]
Georgian Grown
Blueberries
HT-29 colon cancer cells Induce apoptosis
decrease GST
activities
[
39 ]
9.3 Anthocyanins and Cancer
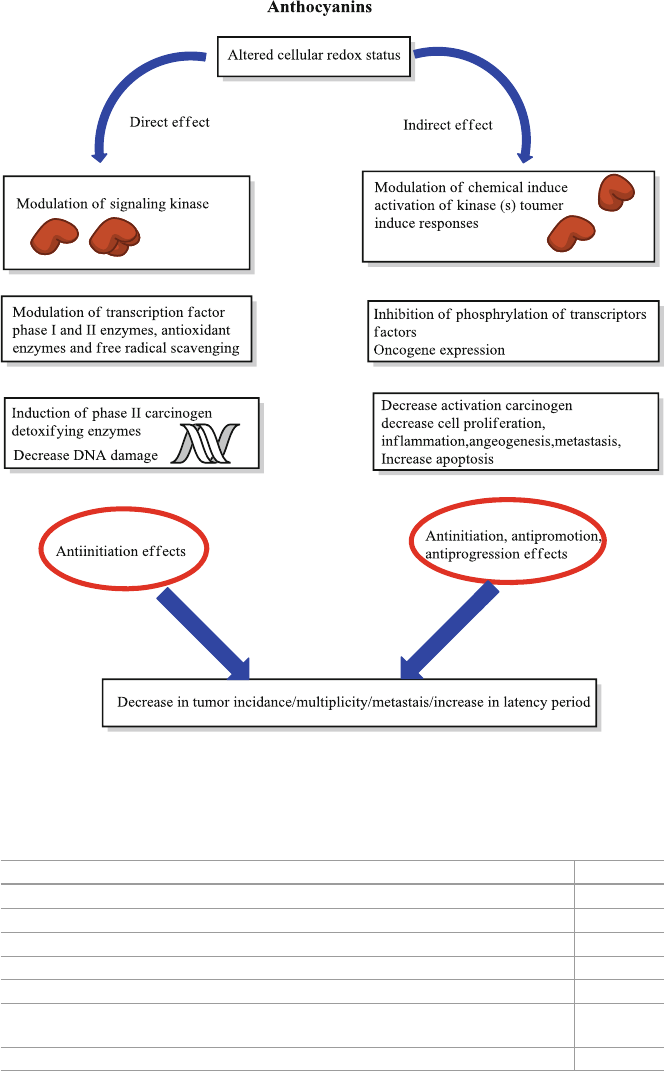
130
Fig. 9.1 Chemopreventive effect of anthocyanins with mechanism
Table 9.2 Mechanisms of anticancer activity of anthocyanins
Mechanism Reference
Antimutagenic activity [
44 ]
Inhibition of oxidative DNA damage [
45 ]
Induction of phase-II enzymes for detoxifi cation and activation of carcinogen [
46 ]
Cellcycle arrest [
47 ]
Induction of apoptosis; inhibition of COX-2 enzymes [
28 ]
Antiangiogenesis e.g. inhibition of H
2
O
2
and tumor necrosis factor α (TNF-α)-
induced VEFG-expression, inhibition of VEGF and VEGF-receptor expression
[ 48 ]
Increased glutathione S transferase (GST) activity [
49 ]
9 Anthocyanins Effects on Carcinogenesis, Immune System and the Central Nervous…
131
9.4 Anthocyanins, Alzheimer Diseases and Brain Function
Anthocyanins prevent Alzheimer’s diseases and neuro-degeneration by increasing
neuronal signaling in brain, mediating memory function and improving glucose
utilization [ 59 ]. Anthocyanins have very benefi cial effects on memory, cognition
and motor function because of delaying the deterioration of neural function [ 59 ,
60 ]. Goyarzu et al. (2004) found in Fisher rats, that blueberry intake suppressed an
aging-associated increase in nuclear factor-kappa B levels (NF-κB) [ 61 ] while
upregulated TNF-α, IL-1β and NF-κB expression in the hippocampus [ 62 ]. Williams
et al. [ 63 ] (2008) demonstrated that blueberry intake induced activation of cyclic
AMP-response element-bonding protein and an increase in the level of brain-
derived neurotrophic factor. Improvement of cerebrovascular blood fl ow may also
be implicated in the favorable effects of berries on brain function [ 64 ]. Ameloriative
effects of anthocyanins in neurodegenerative processes of Alzheimer’s and
Parkinson’s disease is chiefl y due to their ability to decrease infl ammation and oxi-
dative stress in brain. It was observed that berries enhance release of dopamine in
human brain leading to improved communication between brain cells. More con-
sumption of anthocyanin-rich diets like vegetables and fruits and can prevent or
reverse age-related changes in brain and behaviour [ 65 ]. Berries not only stop but
also reverse oxidative-stress-induced and age-related deterioration in neural func-
tion increasing dopamine release thus improving the ability of neurons to increase
intracellular communication [ 66 , 67 ]. Bilberry-fed animals exhibited improved
memory and control of sensory inputs than control animals [ 66 ] while rats given
lyophilized berries indicated improved working memory and increased short-term
memory [ 68 ]. In a brain injury mice-model with middle cerebral artery occlusion
showed neuroprotective effects when fed mulberry fruit extract containing cyanidin-
3- glucoside [ 69 ] (Table 9.3 ).
9.5 Miscellaneous Activities
Anthocyanins can express their antimicrobial activity by causing morphological
damages to bacterial cells or by destroying the structural integrity of the wall,
membrane, intracellular matrix or by cell deformation [ 71 , 72 ]. Four anthocya-
nins namely cyanidin chloride, delphinidin chloride, pelargonidin chloride and
cyanidin- 3- glucoside possess strong anti-bacterial activity against E. coli (gram
−ve) strain CM-871 (DNA repair-defi cient strain) but did not inhibit normal
Escherichia coli and benefi cial probiotic bacteria (gram +ve). That shows the
action association with DNA. In another antibacterial study, anthocyanin exhib-
ited more potential in decreasing the viability of Salmonella enterica var.
typhimurium than other berries phenolic fractions [ 73 ]. The effect was due to
anthocyanins induction of lipopolysaccharide molecules release from the outer
membrane of the gram-negative bacteria.
9.5 Miscellaneous Activities
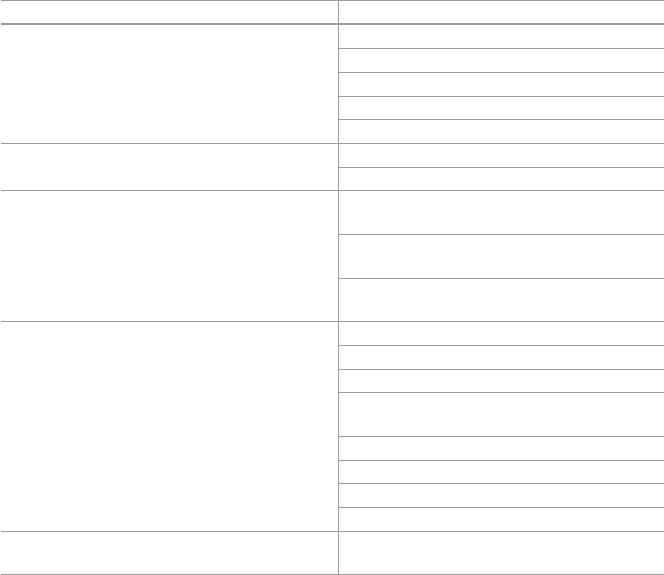
132
Blueberry and billberry extracts showed growth-inhibitory effects on gram nega-
tive bacteria ( Escherchia coli , Enterococcus faecali , Pseudomonas aeuruginosa ,
Citrobacter freundi and Salmonella enterica ser. Typhimurium ) and gram positive
bacteria ( Staphylococusaureus , Listeria monocytogenes , Enterococcus faecalis and
Bacillus subtilis ). Of all bacteria, E. faecalis and C. fruendii were the most sensitive
and E. coli showed the highest resistance [ 74 ]. American cranberry and European
cranberry showed similar anti-bacterial effects [ 75 , 76 ]. Cranberries decreased
growth of E. coli below detectable limits when treated with anthocyanins at native
pH. Anthocyanins exhibited low antimicrobial activity at neutral pH, probably due
to their instability [ 71 ]. Cranberry extract showed no effect on yeast. Blackcurrent
concentrates arrested the growth of E. faecium and S. aureus strains while minor
effects were observed against E. coli [ 77 ]. In another comparative study, microbial
strains exhibited different susceptibility to berry extracts; the order of antimicrobial
Table 9.3 Mechanisms of action of anthocyanins [ 70 ]
Mechanism Anthocyanins
Oxidative stress Scavenges superoxide radicals
Scavenges hydroxyl radicals
Increases antioxidant enzyme activity
Lowers ROS
Decreases NO production
Cardiomyocytes and the endothelium Improves endothelial dysfunction
Mediates vasodilation
Anti-infl ammatory effects Decreases P-selectin, MCP-1, TNF-α
and IL-10 expression
Decreases VEGF and ICAM-1 expression
on endothelial cells
Decreases VLA-4, CD40 and CD36
expression on monocytes
Metabolic effects Decreases LDL-cholesterol
Increases HDL-cholesterol
Inhibits LDL oxidation
Increases faecal expression of acidic
and neutral sterols
Alters sugar and lipid metabolism
Improves insulin resistance
Decreases NFκB levels
Decreases iNOS and COX
2
expression
Anti-ischemic and cardioprotective effects Decreases the magnitude of ischemia–
reperfusion injury
ROS reactive oxygen species, NO nitric oxide, MCP monocyte chemotactic protein, TNF tumour
necrosis factor, IL interleukin, VEGF vascular endothelial growth factor, ICAM intracellular adhe-
sion molecule, VLA very late antigen, CD cluster of differentiation, LDL low density lipoprotein,
HDL high density lipoprotein, NFκB nuclear factor, iNOS inducible nitric oxide synthase, COX
cyclooxygenase
9 Anthocyanins Effects on Carcinogenesis, Immune System and the Central Nervous…
133
effects observed was cloudberries > raspberries > strawberries [ 73 ]. In another inves-
tigation, berry extracts showed bactericidal effects, inhibiting growth of Helicobacter
pylori . All berries extracts showed >70 % inhibition at 1 % concentration with bill-
berry, blueberry and cranberry, extract showing >90 % inhibition [ 78 ].
Anthocyanins of berry extracts (bilberry, blueberry, strawberry, cranberry, rasp-
berry) have shown protective effects against liver damage [ 79 ], ulcer [ 80 ] and
UV-radiation-induced damages especially cyaniding acts as skin-protective agent [ 81 ].
Several enzymes in vitro may be inhibited by anthocyanins [ 82 ] like aldo-
reductase in the lens of rats can be inhibited by both pelargonidin and delphinidin
[ 83 ]. Anthocyanin-3-monoglycosides (namely delphinidin-, petunidin-and malvi-
din-) extracted from grapes inhibited hexokinase, malate dehydrogenase and glyc-
erol dehydrogenase while increased the activity of glutamic acid decarboxylase and
α -glucan-phosphorylase [ 84 ].
Tobacco bud worm larva growth was reported to be inhibited by cyaniding- 3-
glucoside, which may be a useful biological controlling agent [ 85 ].
9.6 Pharmaceutical Products
Anthocyanin containing diets have been used since long as therapeutic remedy
against various diseases in different parts of the world. For example crude extracts
of Vaccinium myrtillus administered intramuscularly and intravenously or ingested
orally can decrease the fragility and permeability of capillaries of blood system in
humans. A health-promoting product, OptiBerry is made form the synergistic com-
bination of 6 selected extracts of wild blueberry and bilberry ( Vaccinium myrtillus
and Vaccinium corymbosum ), elderberries, strawberries, cranberries and raspber-
ries. This product exhibits very high anti-angiogenic, anticarcinogenic, antioxidant,
antibacterial, and anti-atherosclerotic properties [ 86 ]. OptiBerry shows very low
cytotoxicity and can be used as a dietary supplement and safe food. Anthocyanin
powder of red cabbage, grape and tomatoes is used as natural food additives in soft
drinks and confectionary items. Anthocyanins of red cabbage are stable over a
broader pH range and therefore used as substitutes to synthetic blue colorings for
foods with neutral pH [ 87 ]. Anthocyanins of Hibiscus are utilized in herbal formula-
tions as well as in soft drinks [ 88 ].
9.7 Conclusions
With better understanding of cancer subtypes and their risk factors, human clinical
trials should be designed to obtain maximum benefi ts from dietary anthocyanins.
Research in epigenetics and personalized medicines should be carried out to deci-
pher specifi c anthocyanins compounds and determine dosage that may modulate
risk of various age-related diseases in humans. Taking into account the effi cacy and
9.7 Conclusions
134
pharmacological effects of anthocyanins in humans, it is believed that combining
them with other antioxidants like ascorbic acid will be a useful approach for preven-
tion of various diseases. As they target different signaling pathways providing a
complementary and synergistic protective effects against chronic diseases.
References
1. Ahmed, S., Gul, S., Zia-Ul-Haq, M., Riaz, M., & Moga, M. (2015). Anti-infl ammatory effects
of cyclooxygenase-2 inhibitors in rabbits. Pakistan Journal of Zoology, 47 (1), 209–216.
2. Ahmed, S., Gul, S., Gul, H., Zia-Ul-Haq, M., Ercisli, S., & Jaafar, H. Z. E. (2014). Scientifi c
basis for the use of Cinnamonum tamala in cardiovascular and infl ammatory diseases.
Experimental and Clinical Cardiology, 20 , 784–800.
3. Zia-Ul-Haq, M., Khan, B. A., Landa, P., Kutil, Z., Ahmed, S., Qayum, M., et al. (2011).
Platelet aggregation and anti-infl ammatory effects of garden pea, Desi chickpea and Kabuli
chickpea. Acta Poloniae Pharmaceutica, 69 (4), 707–711.
4. Zia-Ul-Haq, M., Landa, P., Kutil, Z., Qayum, M., & Ahmad, S. (2013). Evaluation of anti-
infl ammatory activity of selected legumes from Pakistan: In vitro inhibition of
Cyclooxygenase-2. Pakistan Journal of Pharmaceutical Sciences, 26 (1), 185–187.
5. Ahmed, S., Gul, S., Zia-Ul-Haq, M., & Stanković, M. S. (2014). Pharmacological basis of the
use of Acorus calamus L. in infl ammatory diseases and underlying signal transduction path-
ways. Latin American and Caribbean Bulletin of Medicinal and Aromatic Plant, 13 (1), 38–46.
6. Ahmed, S., Gul, S., Idris, F., Khan, A. H., Zia-Ul-Haq, M., Jaafar, H. Z. E., et al. (2014). Anti-
infl ammatory and antiplatelet activities of plasma are conserved across twelve Mammalian
species. Molecules, 19 (8), 11385–11394.
7. Gul, S., Ahmed, S., Kifl i, N., Uddin, Q. T., Tahir, N. B., Hussain, A., et al. (2014). Multiple
pathways are responsible for anti-infl ammatory and cardiovascular activities of Hordeum vul-
gare L. Journal of Translational Medicine, 12 (1), 1–8.
8. Khanra, R., Dewanjee, S., Dua, T. K., Sahu, R., Gangopadhyay, M., De Feo, V., et al. (2015).
Abroma augusta L. (Malvaceae) leaf extract attenuates diabetes induced nephropathy and car-
diomyopathy via inhibition of oxidative stress and infl ammatory response. Journal of
Translational Medicine, 13 (1), 1–14.
9. Najm-ur-Rahman, M. R., Khan, A., Zia-Ul-Haq, M., & Dima, L. (2015) Mechanism of anti-
infl ammatory and anti-nociceptive actions of Acacia modesta in animal models . Pakistan
Journal of Zoology, 47 (6), 1723–1730.
10. Wang, H., Nair, M. G., Strasburg, G. M., Chang, Y. C., Booren, A. M., Gray, J. I., et al. (1999).
Antioxidant and antiinfl ammatory activities of anthocyanins and their aglycon, cyanidin, from
tart cherries. Journal of Natural Products, 62 (2), 294–296.
11. Seeram, N. P., Momin, R. A., Nair, M. G., & Bourquin, L. D. (2001). Cyclooxygenase inhibi-
tory and antioxidant cyanidin glycosides in cherries and berries. Phytomedicine, 8 (5),
362–369.
12. Rossi, A., Serraino, I., Dugo, P., Di, P. R., Mondello, L., Genovese, T., et al. (2003). Protective
effects of anthocyanins from blackberry in a rat model of acute lung infl ammation. Free
Radical Research, 37 (8), 891–900.
13. Hämäläinen, J. A. (2007). Processing of sound rise time in children and adults with and with-
out reading problems . Jyväskylä: University of Jyväskylä.
14. Miguel, M. (2011). Anthocyanins: Antioxidant and/or anti-infl ammatory activities. Journal of
Applied Pharmaceutical Science, 1 (6), 7–15.
15. Pergola, G., Maldera, S., Tartagni, M., Pannacciulli, N., Loverro, G., & Giorgino, R. (2006).
Inhibitory effect of obesity on gonadotropin, estradiol, and inhibin B levels in fertile women.
Obesity, 14 (11), 1954–1960.
16. Wang, Q., Tang, X. N., & Yenari, M. A. (2007). The infl ammatory response in stroke. Journal
of Neuroimmunology, 184 (1), 53–68.
9 Anthocyanins Effects on Carcinogenesis, Immune System and the Central Nervous…
135
17. Wang, X., Jiang, Y., Yu-Wen, W., Mou-Tuan, H., Chi-Tang, H., & Qingrong, H. (2008).
Enhancing anti-infl ammation activity of curcumin through O/W nanoemulsions. Food
Chemistry, 108 (2), 419–424.
18. Garcia-Alonso, M., Minihane, A. M., Rimbach, G., Rivas-Gonzalo, J. C., & de Pascual-Teresa,
S. (2009). Red wine anthocyanins are rapidly absorbed in humans and affect monocyte che-
moattractant protein 1 levels and antioxidant capacity of plasma. The Journal of Nutritional
Biochemistry, 20 (7), 521–529.
19. Zaid, H., Rayan, A., Said, O., & Saad, B. (2010). Cancer treatment by Greco-Arab and Islamic
herbal medicine. Open Nutraceuticals Journal, 3 , 203–212.
20. Zaid, H., Silbermann, M., Ben-Arye, E., & Saad, S. (2011). Greco-Arab and Islamic herbal-
derived anticancer modalities: From tradition to molecular mechanisms. Evidence-Based
Complementary and Alternative Medicine, 2012 .
21. Saad, B., & Said, O. (2010). Chapter 10: Tradition and prospective of Greco-Arab and Islamic
herbal medicine. In A. Dasgupta & C. Hammett-Stabler (Eds.), Herbal remedies: Toxicity and
effects on clinical laboratory test results . Hoboken: Wiley.
22. Malik, M., Zhao, C., Schoene, N., Guisti, M. M., Moyer, M. P., & Magnuson, B. A. (2003).
Anthocyanin-rich extract from Aronia meloncarpa E. induces a cell cycle block in colon can-
cer but not normal colonic cells. Nutrition and Cancer, 46 (2), 186–196.
23. Kamei, H., Kojima, T., Hasegawa, M., Koide, T., Umeda, T., Yukawa, T., et al. (1995).
Suppression of tumor cell growth by anthocyanins in vitro. Cancer Investigation, 13 (6),
590–594.
24. Kamei, H., Hashimoto, Y., Koide, T., Kojima, T., & Hasegawa, M. (1998). Anti-tumor effect
of methanol extracts from red and white wines. Cancer Biotherapy and Radiopharmaceuticals,
13 (6), 447–452.
25. Zhao, L., Gu, J., Dong, A., Zhang, Y., Zhong, L., He, L., et al. (2005). Potent antitumor activity
of oncolytic adenovirus expressing mda-7/IL-24 for colorectal cancer. Human Gene Therapy,
16 (7), 845–858.
26. Zhang, X., Yeeleng, Y., Dong, W., Gu, C., & Feng, C. (2008). Novel omics technologies in
nutrition research. Biotechnology Advances, 26 (2), 169–176.
27. Jing, P., Bomser, J. A., Schwartz, S. J., He, J., Magnuson, B. A., & Giusti, M. M. (2008).
Structure- function relationships of anthocyanins from various anthocyanin-rich extracts on
the inhibition of colon cancer cell growth. Journal of Agricultural and Food Chemistry, 56 (20),
9391–9398.
28. Yi, W., Fischer, J., & Akoh, C. C. (2005). Study of anticancer activities of muscadine grape
phenolics in vitro. Journal of Agricultural and Food Chemistry, 53 (22), 8804–8812.
29. Seeram, N. P., Adams, L. S., Henning, S. M., Niu, Y., Zhang, Y., Nair, M. G., et al. (2005). In
vitro antiproliferative, apoptotic and antioxidant activities of punicalagin, ellagic acid and a
total pomegranate tannin extract are enhanced in combination with other polyphenols as found
in pomegranate juice. Journal of Nutrition and Biochemistry, 16 (6), 360–367.
30. Lala, G., Malik, M., Zhao, C., He, J., Kwon, Y., Giusti, M. M., et al. (2006). Anthocyanin-rich
extracts inhibit multiple biomarkers of colon cancer in rats. Nutrition and Cancer, 54 (1),
84–93.
31. Cvorovic, J., Tramer, F., Granzotto, M., Candussio, L., Decorti, G., & Passamonti, S. (2010).
Oxidative stress-based cytotoxicity of delphinidin and cyanidin in colon cancer cells. Archives
of Biochemistry and Biophysics, 501 (1), 151–157.
32. Kang, S. Y., Seeram, N. P., Nair, M. G., & Bourquin, L. D. (2003). Tart cherry anthocyanins
inhibit tumor development in Apc sup Min/sup mice and reduce proliferation of human colon
cancer cells. Cancer Letters, 194 (1), 13–19.
33. Zu, X. Y., Zhang, Z. Y., Zhang, X. W., Yoshioka, M., Yang, Y. N., & Li, J. (2010). Anthocyanins
extracted from Chinese blueberry (Vaccinium uliginosum L.) and its anticancer effects on
DLD-1 and COLO205 cells. Chinese Medical Journal, 123 (19), 2714.
34. Cai, H., Marczylo, T. H., Teller, N., Brown, K., Steward, W. P., Marko, D., et al. (2010).
Anthocyanin-rich red grape extract impedes adenoma development in the ApcMin mouse:
Pharmacodynamic changes and anthocyanin levels in the murine biophase. European Journal
of Cancer, 46 (4), 811–817.
References
136
35. Misikangas, M., Pajari, A. M., Päivärinta, E., Oikarinen, S. I., Rajakangas, J., Marttinen, M.,
et al. (2007). Three Nordic berries inhibit intestinal tumorigenesis in multiple intestinal neo-
plasia/+ mice by modulating β-catenin signaling in the tumor and transcription in the mucosa.
Journal of Nutrition, 137 (10), 2285–2290.
36. Singletary, K. W., Stansbury, M. J., Giusti, M., Van Breemen, R. B., Wallig, M., & Rimando,
A. (2003). Inhibition of rat mammary tumorigenesis by concord grape juice constituents.
Journal of Agricultural and Food Chemistry, 51 (25), 7280–7286.
37. Stoner, G. D., Li-Shu, W., Zikri, N., Chen, T., Stephen, S. H., Chuanshu, H., et al. (2007).
Cancer prevention with freeze-dried berries and berry components. Seminars in Cancer
Biology, 17 , 403–410.
38. Shin, D. Y., Lu, J. N., Kim, G. Y., Jung, J. M., Kang, H. S., Lee, W. S., et al. (2011). Anti-
invasive activities of anthocyanins through modulation of tight junctions and suppression of
matrix metalloproteinase activities in HCT-116 human colon carcinoma cells. Oncology
Reports, 25 (2), 567.
39. Srivastava, A., Akoh, C. C., Fischer, J., & Krewer, G. (2007). Effect of anthocyanin fractions
from selected cultivars of Georgia-grown blueberries on apoptosis and phase II enzymes.
Journal of Agricultural and Food Chemistry, 55 (8), 3180–3185.
40. Zhang, Y., Vareed, S. K., & Nair, M. G. (2005). Human tumor cell growth inhibition by non-
toxic anthocyanidins, the pigments in fruits and vegetables. Life Sciences, 76 (13),
1465–1472.
41. Stoner, G. D., Kresty, L. A., Carlton, P. S., Siglin, J. C., & Morse, M. A. (1999). Isothiocyanates
and freeze-dried strawberries as inhibitors of esophageal cancer. Toxicological Sciences,
52 (suppl 1), 95–100.
42. He, J., Magnuson, B. A., & Giusti, M. M. (2005). Analysis of anthocyanins in rat intestinal
contents impact of anthocyanin chemical structure on fecal excretion. Journal of Agricultural
and Food Chemistry, 53 (8), 2859–2866.
43. Carlton, P. S., Kresty, L. A., & Stoner, G. D. (2000). Failure of dietary lyophilized strawberries
to inhibit 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone-and benzo [a] pyrene-induced lung
tumorigenesis in strain A/J mice. Cancer Letters, 159 (2), 113–117.
44. Ohara, A., & Matsuhisa, T. (2004). Effects of diet composition on mutagenic activity in urine.
BioFactors, 22 (1), 115–118.
45. Singletary, K. W., Jung, K.-J., & Giusti, M. (2007). Anthocyanin-rich grape extract blocks
breast cell DNA damage. Journal of Medicinal Food, 10 (2), 244–251.
46. Shih, P.-H., & Yen, G.-C. (2007). Differential expressions of antioxidant status in aging rats: The
role of transcriptional factor Nrf2 and MAPK signaling pathway. Biogerontology, 8 (2), 71–80.
47. Renis, M., Calandra, L., Scifo, C., Tomasello, B., Cardile, V., Vanella, L., et al. (2008).
Response of cell cycle/stress-related protein expression and DNA damage upon treatment of
CaCo
2
cells with anthocyanins. British Journal of Nutrition, 100 (01), 27–35.
48. Bagchi, A., Papazoglu, C., Wu, Y., Capurso, D., Brodt, M., Francis, D., et al. (2007). CHD5 is
a tumor suppressor at human 1p36i. Cell, 128 (3), 459–475.
49. Boateng, J., Verghese, M., Shackelford, L., Walker, L. T., Khatiwada, J., Ogutu, S., et al.
(2007). Selected fruits reduce azoxymethane (AOM)-induced aberrant crypt foci (ACF) in
Fisher 344 male rats. Food and Chemical Toxicology, 45 (5), 725–732.
50. Tramer, F., Moze, S., Ademosun, A. O., Passamonti, S., & Cvorovic, J. Dietary anthocyanins:
Impact on colorectal cancer and mechanisms of action . In R. Ettarh (Ed.) Colorectal Cancer
- From Prevention to Patient Care. InTech,Serbia.
51. Katsube, N., Iwashita, K., Tsushida, T., Yamaki, K., & Kobori, M. (2003). Induction of apop-
tosis in cancer cells by bilberry (Vaccinium myrtillus) and the anthocyanins. Journal of
Agricultural and Food Chemistry, 51 (1), 68–75.
52. Johnson, S. M., Wang, X., & Evers, B. M. (2011). Triptolide inhibits proliferation and migra-
tion of colon cancer cells by inhibition of cell cycle regulators and cytokine receptors. Journal
of Surgical Research, 168 (2), 197–205.
53. Thomasset, S., Teller, N., Cai, H., Marko, D., Berry, D. P., Steward, W. P., et al. (2009). Do
anthocyanins and anthocyanidins, cancer chemopreventive pigments in the diet, merit develop-
ment as potential drugs? Cancer Chemotheraphy and Pharmacology, 64 (1), 201–211.
9 Anthocyanins Effects on Carcinogenesis, Immune System and the Central Nervous…
137
54. Rossi, M., Garavello, W., Talamini, R., Negri, E., Bosetti, C., Dal, M. L., et al. (2007).
Flavonoids and the risk of oral and pharyngeal cancer: A case-control study from Italy. Cancer
Epidemiology, Biomarkers and Prevention, 16 (8), 1621–1625.
55. Bosetti, C., Gallus, S., & La Vecchia, C. (2006). Aspirin and cancer risk: An updated quantita-
tive review to 2005. Cancer Causes and Control, 17 (7), 871–888.
56. Stoner, G. D. (2009). Foodstuffs for preventing cancer: The preclinical and clinical develop-
ment of berries. Cancer Prevention Research, 2 (3), 187–194.
57. Huang, B., Zhao, J., Unkeless, J. C., Feng, Z. H., & Xiong, H. (2008). TLR signaling by tumor
and immune cells: A double-edged sword. Oncogene, 27 (2), 218–224.
58. Nagase, T., Ishikawa, K., Suyama, M., Kikuno, R., Hirosawa, M., & Miyajima, N. (1998).
Prediction of the coding sequences of unidentifi ed human genes. XII. The complete sequences
of 100 new cDNA clones from brain which code for large proteins in vitro. DNA Research,
5 (6), 355–364.
59. Krikorian, R., Eliassen, J. C., Boespfl ug, E. L., Nash, T. A., & Shidler, M. D. (2010). Improved
cognitive-cerebral function in older adults with chromium supplementation. Nutritional
Neuroscience, 13 (3), 116–122.
60. Krikorian, R., Nash, T. A., Shidler, M. D., Shukitt-Hale, B., & Joseph, J. A. (2010). Concord
grape juice supplementation improves memory function in older adults with mild cognitive
impairment. British Journal of Nutrition, 103 (05), 730–734.
61. Goyarzu, P., Malin, D. H., Lau, F. C., Taglialatela, G., Moon, W. D., Jennings, R., et al. (2004).
Blueberry supplemented diet: Effects on object recognition memory and nuclear factor-kappa
B levels in aged rats. Nutritional Neuroscience, 7 (2), 75–83.
62. Shukitt-Hale, B., Lau, F. C., Carey, A. N., Galli, R. L., Spangler, E. L., Ingram, D. K., et al.
(2008). Blueberry polyphenols attenuate kainic acid-induced decrements in cognition and alter
infl ammatory gene expression in rat hippocampus. Nutritional Neuroscience, 11 (4),
172–182.
63. Williams, C. M., El Mohsen, M. A., Vauzour, D., Rendeiro, C., Butler, L. T., Ellis, J. A., et al.
(2008). Blueberry-induced changes in spatial working memory correlate with changes in hip-
pocampal CREB phosphorylation and brain-derived neurotrophic factor (BDNF) levels. Free
Radical Biology and Medicine, 45 (3), 295–305.
64. Spencer, J. P. (2010). The impact of fruit fl avonoids on memory and cognition. British Journal
of Nutrition, 104 (S3), S40–S47.
65. Lau, F. C., Bielinski, D. F., & Joseph, J. A. (2007). Inhibitory effects of blueberry extract on
the production of infl ammatory mediators in lipopolysaccharide activated BV2 microglia.
Journal of Neuroscience Research, 85 (5), 1010–1017.
66. Galli, R. L., Bielinski, D. F., Szprengiel, A., Shukitt-Hale, B., & Joseph, J. A. (2006). Blueberry
supplemented diet reverses age-related decline in hippocampal HSP70 neuroprotection.
Neurobiology of Aging, 27 (2), 344–350.
67. Youdim, K. A., Martin, A., & Joseph, J. A. (2000). Incorporation of the elderberry anthocya-
nins by endothelial cells increases protection against oxidative stress. Free Radical Biology
and Medicine, 29 (1), 51–60.
68. Ramirez, M. R., Izquierdo, I., do Carmo, B. R. M., Zuanazzi, J. A., Barros, D., & Henriques,
A. T. (2005). Effect of lyophilised Vaccinium berries on memory, anxiety and locomotion in
adult rats. Pharmacological Research, 52 (6), 457–462.
69. Kang, T. H., Hur, J. Y., Kim, H. B., Ryu, J. H., & Kim, S. Y. (2006). Neuroprotective effects of
the cyanidin-3-O-β-d-glucopyranoside isolated from mulberry fruit against cerebral ischemia.
Neuroscience Letters, 391 (3), 122–126.
70. Kruger, D. J., Greenberg, E., Murphy, J. B., DiFazio, L. A., & Youra, K. R. (2014). Local
concentration of fast-food outlets is associated with poor nutrition and obesity. American
Journal of Health Promotion, 28 (5), 340–343.
71. Lacombe, A., Wu, V. C., Tyler, S., & Edwards, K. (2010). Antimicrobial action of the American
cranberry constituents; phenolics, anthocyanins, and organic acids, against Escherichia coli
O157: H7. International Journal of Food Microbiology, 139 (1), 102–107.
References
138
72. Cisowska, A., Wojnicz, D., & Hendrich, A. B. (2011). Anthocyanins as antimicrobial agents
of natural plant origin. Natural Product Communications, 6 (1), 149–156.
73. Nohynek, L. J., Alakomi, H. L., Kähkönen, M. P., Heinonen, M., Helander, I. M., Oksman-
Caldentey, K. M., et al. (2006). Berry phenolics: Antimicrobial properties and mechanisms of
action against severe human pathogens. Nutrition and Cancer, 54 (1), 18–32.
74. Burdulis, D., Sarkinas, A., Jasutiené, I., Stackevicené, E., Nikolajevas, L., & Janulis, V. (2008).
Comparative study of anthocyanin composition, antimicrobial and antioxidant activity in bil-
berry (Vaccinium myrtillus L.) and blueberry (Vaccinium corymbosum L.) fruits. Acta Poloniae
Pharmaceutica, 66 (4), 399–408.
75. Cesoniene, L., Jasutiene, I., & Sarkinas, A. (2008). Phenolics and anthocyanins in berries of
European cranberry and their antimicrobial activity. Medicina, 45 (12), 992–999.
76. Wu, V. C.-H., Xujian, Q., Alfred, B., & Laura, H. (2008). Antibacterial effects of American
cranberry (Vaccinium macrocarpon) concentrate on foodborne pathogens. LWT--Food Science
and Technology, 41 (10), 1834–1841.
77. Werlein, C. H.-D., Kütemeyer, G., Schatton, E. M., & Hubbermann, K. S. (2005). Infl uence of
elderberry and blackcurrant concentrates on the growth of microorganisms. Food Control,
16 (8), 729–733.
78. Chatterjee, A., Yasmin, T., Bagchi, D., & Stohs, S. J. (2004). Inhibition of Helicobacter pylori
in vitro by various berry extracts, with enhanced susceptibility to clarithromycin. Molecular
and Cellular Biochemistry, 265 (1-2), 19–26.
79. Wang, S. Y., & Lin, H.-S. (2000). Antioxidant activity in fruits and leaves of blackberry, rasp-
berry, and strawberry varies with cultivar and developmental stage. Journal of Agricultural
and Food Chemistry, 48 (2), 140–146.
80. Kong, J. M., Chia, L. S., Goh, N. K., Chia, T. F., & Brouillard, R. (2003). Analysis and biologi-
cal activities of anthocyanins. Phytochemistry, 64 (5), 923–933.
81. Sharma, R. (2001). Impact of solar UV-B on tropical ecosystems and agriculture. Case study:
Effect of UV-B on rice. Proceedings of Seawpit98 and Seawpit2000, 1 , 92–101.
82. Galvez, J., De La Cruz, J. P., Zarzuelo, A., Sanchez, D. L., & Cuesta, F. (1995). Flavonoid
inhibition of enzymic and nonenzymic lipid peroxidation in rat liver differs from its infl uence
on the glutathione-related enzymes. Pharmacology, 51 (2), 127–133.
83. Varma, S. D., & Kinoshita, J. H. (1976). Inhibition of lens aldose reductase by fl avonoids—
Their possible role in the prevention of diabetic cataracts. Biochemical Pharmacology, 25 (22),
2505–2513.
84. Beltoft, V. M., Binderup, M.-L., Frandsen, H. L., Lund, P., & Nørby, K. K. (2013). EFSA cef
panel (EFSA panel on food contact materials, enzymes, fl avourings and processing aids),
2013. scientifi c opinion on fl avouring group evaluation 21, revision 4 (fge. 21rev4) . European
Food Safety Authority.
85. Delgado-Vargas, F., Jiménez, A., & Paredes-López, O. (2000). Natural pigments: carotenoids,
anthocyanins, and betalains—Characteristics, biosynthesis, processing, and stability. Critical
Reviews in Food Science and Nutrition, 40 (3), 173–289.
86. Zafra-Stone, S., Yasmin, T., Bagchi, M., Chatterjee, A., Vinson, J. A., & Bagchi, D. (2007).
Berry anthocyanins as novel antioxidants in human health and disease prevention. Molecular
Nutrition and Food Research, 51 (6), 675–683.
87. Bridle, P., & Timberlake, C. (1997). Anthocyanins as natural food colours-selected aspects.
Food Chemistry, 58 (1), 103–109.
88. Oancea, S., & Oprean, L. (2011). Anthocyanin extracts in the perspective of health benefi ts
and food applications. Revista de Economia, 218 .
9 Anthocyanins Effects on Carcinogenesis, Immune System and the Central Nervous…
