
Composing Popular Music with Physarum
polycephalum-based Memristors
Satvik Venkatesh
Interdisciplinary Centre for
Computer Music Research
University of Plymouth
United Kingdom
satvik.venkatesh
@plymouth.ac.uk
Edward Braund
Interdisciplinary Centre for
Computer Music Research
University of Plymouth
United Kingdom
edward.braund
@plymouth.ac.uk
Eduardo Reck Miranda
Interdisciplinary Centre for
Computer Music Research
University of Plymouth
United Kingdom
eduardo.miranda
@plymouth.ac.uk
ABSTRACT
Creative systems such as algorithmic comp osers often use
Artificial Intelligence models like Markov chains, Neural
Networks, and Genetic Algorithms to model stochastic pro-
cesses. Unconventional Computing (UC) technologies ex-
plore non-digital ways of data storage, processing, input,
and output. UC paradigms such as Biocomputing and Quan-
tum Computing delve into domains beyond the binary bit
to handle complex non-linear functions. In this pap er, we
harness Physarum polycephalum as a memristor to process
and generate creative data for p opular music. The organism
works as a collaborator in the process of composing our song
titled Creep into my Lawn. While there has been research
conducted in this area, the literature lacks examples of pop-
ular music and how the organism’s non-linear behaviour can
b e controlled while composing music. This is important be-
cause non-linear forms of representation are not as obvious
as conventional digital means. This study aims at dissemi-
nating this technology to non-experts and musicians so that
they can incorporate it in their creative processes. Further-
more, it combines resistors and memristors to have more
flexibility while generating music and optimises parameters
for faster processing and performance.
Author Keywords
Unconventional Computing, Biocomputing, Musical Com-
p osition, Popular Music, Physarum polycephalum
CCS Concepts
•Theory of computation → Unconventional Com-
puting; •Applied computing → Sound and Music
Computing; Algorithmic Composition;
1. INTRODUCTION
There is a vast corpus of research that explores computa-
tional processes to exhibit creative behaviour [8]. The pro-
cess of musical composition frequently involves two elements
— material that has been previously heard and ideas that
are new and innovative [9]. The former encompasses char-
acteristics of the composer’s style and their inspirations.
The latter has been explored in computational creativity
Licensed under a Creative Commons Attribution
4.0 International License (CC BY 4.0). Copyright
remains with the author(s).
NIME’20, July 21-25, 2020, Royal Birmingham Conservatoire,
Birmingham City University, Birmingham, United Kingdom.
through stochastic processes, which aim to induce random-
ness in the output of the system [19].
Many Artificial Intelligence (AI) models such as Markov
chains [12], Genetic Algorithms (GA), Artificial Neural Net-
works (ANNs) [23] have been successfully adopted to ren-
der creative systems. Markov chains compute a conditional
probability distribution of future states depending on the
current state [19]. The architecture of ANNs is inspired by
the structure of the brain and enables us to model com-
plex non-linear functions. Additionally, other mode ls such
as self-similarity and cellular automata [14], that do not
strictly fall under the AI bracket, have been implemented
to render creative systems [10].
Physical, chemical, and biological processes have not only
inspired AI algorithms, but given rise to other forms of com-
putation. Unconventional C omputing (UC) aims to develop
new computer architectures for data storage and processing
[1]. Computational creativity has been conventionally ren-
dered on classical computers based on the von Neumann
architecture. The fundamental unit of processing and stor-
age is the binary bit, that is either 0 or 1. Conversely, UC
technologies explore analog forms of computation and stor-
age. For example, quantum computing uses qubits as the
fundamental unit of processing.
Physarum polycephalum is a plasmodial slime mould that
has been widely harnessed for biocomputing applications.
Researchers have utilised its ability to optimise its shape ac-
cording to the environment to solve mazes [16], build logic
gates [2], and find shortest paths [17]. Another interesting
prop erty of the organism is that it can act as a memristor
[11]. Memristor (a portmanteau of memory and resistor)
is the fourth fundamental passive circuit element, the first
three being resistor, capacitor, and inductor. It was theo-
retically demonstrated by Leon Chua in 1971 [7], but was
physically realised in 2008 [22].
Research i nto memristor technology is gaining traction
due to many reasons. It combines storage and process-
ing into one unit and therefore, has the potential to opti-
mise computing architectures. [21, 13] demonstrated spike-
timing-dep endent plasticity (STDP)-like behaviour using
memristors. STDP has been adopted in feed-forward ANNs
for pattern recognition algorithms [3].
Physarum polycephalum-based memristors serve as an in-
expens ive and accessible UC prototype, when compared to
quantum computers and chemically manufactured memris-
tors. [5] designed 3D printed receptacles to enclose the or-
ganism into a portable electrical component. [6] presented a
compact kernel to interface with biomemristors for creative
practitioners and D.I.Y. prototyping enthusiasts. Moreover,
research is progressing with regards to making UC tech-
nologies available to individuals outside laboratories. This
514

makes it increasingly important to use these bespoke sys-
tems in creative works.
This pap er explores how Physarum polycephalum-based
memristors can be used in the process of composing popular
music. It works with Physarum polycephalum as a collabo-
rator and is a step towards the long-term dream o f building
a rock band of biomemristors. As we are using a non-linear
comp onent as the processing device, input parameters that
govern the composition process require non-digital forms
of representation. This study renders a practice-based ap-
proach towards understanding non-linear input param eters,
which m ay not be as obvious as binary logic. It aims to
break down the creative process and explores how these pa-
rameters influence the output of the PhyBox. Biocomputer
Rhythms by Eduardo Miranda was the first musical piece
for piano and percussion composed with the PhyBox [15].
However, it was not composed in a popular music context
and there have been recent developments with mapping pro-
cedures, such as resistors can be used alongside memristors
in the process of composition [24]. This paper incorporates
these developments, optimises parameters to minimise pro-
cessing time, and offers more degree of freedom to the user.
2. SYSTEM OVERVIEW
2.1 Hardware-wetware Framework
We maintain a Physarum polycephalum farm [1] grown in
plastic lunch boxes as shown in figure 1. In order to grow
Physarum polycephalum-based memristors and make them
a part of the composition process, we transfer a section of
the organism into receptacles [5]. These receptacles allow
the organism to act as an electrical component and are oper-
ated through the PhyBox [6], a portable kernel for creative
applications as shown in figure 2. PhyBox is powered by a
Raspbe rry Pi model B.
Figure 1: Physarum polycephalum culture grown in
a lunch box.
Training data of music is converted into voltages with the
help of a mapping procedure [24]. In this paper, training
data refers to the music originally composed by the author.
These voltages are serially applied across the memristors as
shown in figure 3. The current flowing through the memris-
tor depends on its resistance (or memristance). The current
is again translated to music through a similar mapping pro-
cedure. Therefore, the memristors create new music based
on the training data.
Music is fed into the PhyBox through the MIDI proto-
col. MIDI data is broken down into four parameters —
pitch, velocity, time between note-ons (rhythm), and dura-
Figure 2: Setting up the PhyBox for musical com-
position.
tion. Each memristor processes one of these parameters in
parallel, that is there are 4 memristors in the circuit.
2.2 Mapping procedure
The framework for generating creative data harnesses the
spiking behaviour of memristors. Each time there i s a volt-
age change across the memristor, there is a spike in current
as shown in figure 3. This behaviour is different from that of
linear components like resistors in electronic circuits. The
following explains its behaviour. When the positive change
in voltage is produced across the memristor, the resistance
is initially low. With time, it develops resistance and there
is a gradual decrease in the current flowing through the cir-
cuit.
Figure 3: Spike generated by the memristor on ap-
plying four voltage changes — 0.75V, 1.5V, 0.5V,
and 1.5V. Positive spikes are produced for positive
voltage changes and vice versa.
2.2.1 Music to Voltages
In this system, these voltage transitions are attributed to
musical note transitions. A musical note is assigned to a
sp ecific voltage on a priority basis. Musical notes that occur
more often are assigned higher priority and vice versa. If
there are two events with the same number of occurrences,
the more recent event is given a higher priority. This is
analogous to the learning rule in STDP, where the new event
is good and the old event is bad.
As we are using a Raspberry Pi, the voltage range is 0 to
3V. It is assigned by equation 1 [24].
515

v
a
=
3
N
(p −
1
2
)V olts (1)
where v
a
is the assigned voltage, N is the numb er of musical
notes and p is the priority of the event. A relationship
b etween popularity and assigned voltages is devised by the
following analogy. The rate of change of current (or slope)
in the beginning of the graph is greater when compared to
the end. Moreover, taller spikes are generated for greater
changes in voltage. The stability of resistance is related to
the popularity of the e vent. Therefore, more popular events
are mapped to lower voltages and less popular events are
mapp ed to higher voltages.
Note that when the training data is initially fed into the
PhyBox, there are no events in the memory. It is trained
during the course of the song. Therefore, the number of
variations that can possibly occur gradually increases with
more data.
2.2.2 Current to Music
A similar procedure is followed while converting current
readings to music. The voltage range is well-defined to be 0
to 3V. However, a discrete current range is not defined be-
cause the resistance of the memristor is unknown. Empiri-
cally, studies have approximated the resistance in the order
of mega-ohms [6]. Let I
max
and I
min
b e the maximum and
minimum current sensed during the spike respectively. The
current range [I
1
, I
2
] is defined by the following equation.
[I
1
, I
2
] =
h
min(V
a
)
(v
a
/I
min
)
,
max(V
a
)
(v
a
/I
max
)
i
(2)
where V
a
is the set of all assigned voltages and v
a
the input
voltage at that specific moment in time. The current range
is di vided into equal divisions and converted to music based
on midrise quantisation [4]. Again, the events are ordered
according to priority, where lower current values stand for
more popular events and vice versa.
2.2.3 Resistors
One of the advantages of this mapping procedure is that
it provides a way to incorporate resistors in the process of
musical composition. It defines a resistor’s behaviour as lin-
ear. Therefore, when the training data is fed into a resistor,
the output of the system is exactly identical to the input.
This offers creative pathways to explore in the process of
comp osition, such as the rhythm can be identical to the in-
put and the pitch can be composed by the memristor. It is
imp ortant to note that the value of the resistance does not
influence the output. In this study, we used 330kΩ resistors.
2.3 Parameters
There are two parameters that monitor the output produced
by the system — dwell time and measurement offset. Dwell
time specifies how long the voltage is applied across the
memristor. This basically determines the processing time
of the memristors for each musical event. For example, if
the dwell time is 1 s, the memristor takes 20 s to process
20 musical events. The measurement offset determines at
what point of time is the current value calculated in the
circuit. For instance, on applying 1.5 V at 0.1 s in figure
3, 0% offset considers the current at 0.1 s and 100% offset
considers the current at 1.1 s (Here the dwell time was 1
s). [6, 24] have conducted some experiments to understand
how these non-linear parameters influence the output of the
memristors. 0% offset produces smaller deviations from the
input data when com pared to 100% offset [24]. However,
there is still not a clear delineation of these functions due
to their complex non-linear behaviour.
3. EXPERIMENTS
In this paper, we us e Physarum polycephalum as a collabo-
rator in the process of musical comp osition. As the current
interface only accepts MIDI data, we have harnessed it to
comp ose parts for piano, drums, and the vocal melody. The
dwell time was constant at 1 s. The measurement offset was
altered between 0%, 30%, 50%, and 100%.
3.1 Structure
The time and key signature of the song is 4/4 and E mi-
nor respectively. The genre of the song falls under Rock.
The song starts with the main riff played only on the pi-
ano, which converges into the other instruments joining the
main riff. Followed by that, we have one verse, pre-chorus,
and chorus, which are sung by a vocalist. The chorus has
the main riff in the background. After the chorus, there
is a piano solo followed by a drum solo. The song fin-
ishes with another chorus at the end. This song follows
the structure adopted by most popular music compositions.
However, it was slightly altered, for instance omitting the
second verse, in order to make things more interesting in
the view of Physarum polycephalum.
In order to explain the structure of the song with clar-
ity, the verse was divided into two parts. While feeding in
data for piano and drums, we divided the whole song into
3 blocks. This was done to avoid overloading the model
with too m any possibilities and transitions. The first block
consisted of the intro and first part of the verse. The sec-
ond block comprised the second part of the verse and the
pre-chorus. The third block contained the remainder of the
song. These blocks were not divided with regards to time,
but were divided with respect to number of MIDI events
and structure of the song. For instance, chords in the verse
contained more MIDI events than melody lines. The vocal
melo dy was transcribed as a MIDI file. MIDI events like
sustain pedal was removed from the training data.
3.2 Main Riff
The main riff of the song is 4 bars in length. A piano tune
is backed up by drums and guitars for the main riff. The
first two bars of the piano melody were composed by the
author. We wanted the third and the fourth bar to follow a
similar pattern as the first two, but with minor variations.
The aim is to create a feeling of repetition alongside some
anticipation in the listener. We conducted experiments with
measurement offset values of 0%, 30%, and 100%. The gui-
tars and drums were composed by keeping the piano track
as a reference.
3.3 Verse
The instrumentation for the verse consisted of vocals, pi-
ano, and drums. The first part of the verse had simple pi-
ano chords at the beginning of each bar. We experimented
with measurement offset values of 0%, 30%, and 100%. The
objective of this experiment was to make Physarum poly-
cephalum improve the chord pattern, chord inversions, and
the voicing.
The second part of the verse had arpeggios played on the
piano. For this section of the song, we felt that there was less
flexibility with altering the rhythm. We wanted to maintain
the rhythmic structure originally composed by the author.
Therefore, we attached m emristors for pitch and velocity
and resistors for time between not e-ons and duration. The
measurement offset was set to 100% and this configuration
was refe rred to as 100R.
3.4 Piano Solo
516

The harmonic and rhythmic complexity of the piano solo is
higher than the other sections of the song. The piano solo
was originally composed by the author. Also, there was a
scale shift within the solo from E minor to E Phrygian and
then C major. At the end of the piano solo, the song re-
turns to E minor. We wanted to explore how Physarum
po lycephalum would respond to scale changes and complex
melo dic ideas. This is the most interesting section of the
song because it provides a high degree of freedom with re-
gards to musical options. Additionally, it does not restrict
Physarum polycephalum with respect to the song’s instru-
mentation because the ot her instruments do not play during
the piano solo. Tests were conducted for measurement off-
sets of 5%, 30%, 50%, 100%, and 100%R.
3.5 Drums
We wanted to harness the organism to create new drum
fills based on existing ones. Additionally, we wanted to
explore its creativity for the drum solo. We tested it with
measurement offset values of 0%, 30%, 100%, and 100%R.
3.6 Vocal Tune
A vocal melody was composed by the author and tran-
scribe d as a MIDI track. Our objective of using Physarum
po lycephalum for the vocal tune was to make it sound more
catchy and less monotonous. We explored measurement off-
sets of 0%, 30%, 100%, and 100%R.
3.7 Dissimilarity
In order to objectively calculate the difference between orig-
inal melody and Physarum polycephalum’s rendition, we de-
fined a metric called dissimilarity, which is the number of
musical events that are different from the original input.
Each of the parameters — pitch, velocity, time between
note-ons, and duration would return a value of dissimilarity.
The total dissimilarity is defined as the s um of individual
dissimilarities of the four parameters.
4. RESULTS
All audio and MIDI files that were composed by the au-
thor and Physarum polycephalum can be found in this link
1
.
The naming format for the files is {block
Instrument Type
Offset}. The title of the song is Creep into my Lawn. A mu-
sic video that illustrates the parts composed by Physarum
p olycephalum and the author is available in this link
2
. The
mo del used to generate musical data only uses note transi-
tions. This gives high flexibility to Physarum polycephalum,
but opens up possibilities like time signature mismatch and
disharmonious chords. Therefore, few modifications were
made by the composer to suit the song. As the model is
based on musical transitions, in Physarum polycephalum’s
output, the first note will be missing because no transition
has occurred yet. Hence, while evaluating its output, the
first note was added from the input.
4.1 Dissimilarity
Table 1 contains the total dissimilarities for different values
of measurement offset. Consistently, 0% was more similar
to the input when compared to other offsets. 30% was closer
to the input when compared to 1 00%, but the difference was
much smaller.
4.2 Main Riff
1
https://github.com/satvik-venkatesh/
Physarum-Music
2
https://youtu.be/dKQhnlO7ZrI
The main riff originally composed by the author is shown in
figure 4. Physarum polycephalum’s output for measurement
offset of 0% was not interesting. It resembled the origi-
nal melody except for changing one crotchet to a dotted
crotchet. For measurement offsets of 30% and 100%, the
generated output sounded more appropriate for the main
riff. The authors felt that 30% offset had more resem-
blance to the original tune when compared to 100%. We
chose 100% and made no modifications to Physarum poly-
cephalum’s output. It is shown in figure 5. For the third
and fourth bar of the main riff, guitar and drums parts
were re-composed to match the new tune composed by the
memristors.
Figure 4: The original main riff composed by the
author.
Figure 5: Physarum polycephalum’s rendition of the
main riff for measurement offset of 100%.
4.3 Verse Part 1
The first part of the verse essentially contained piano chords,
drums, and vocals. 0% offset subtly altered the harmonic
structure of the verse. 30% output collapsed all t he indi-
vidual notes into one point. This was because the time
b etween the chords was 0, which was obviously not useful
for the composition. 100% made too many variations that
did not suit the genre of the song and it was difficult to
p erceive the harmonic structure. Hence, we chose 0% offset
for this section.
The original chord pattern of the verse was Em—G—
Am—D. All chords were first inversions. While processing
the output of Physarum polycephalum, the root notes of
chords were left unchanged. The first chord was the same
as the input. The second chord was a triad of the notes
E4, G4, and A4 (the numbers stand for the octave), which
sounded very close to the chord Esus4/G. In order to form a
triad that sounds harmonically more stable, G4 was changed
to C5 and thus, obtaining Esus4/G. The third chord gener-
ated by Physarum polyceph alum contained E2 as the low-
est note, which sounded like the second inversion of Am.
Therefore, the third chord was changed to the second in-
version of Am. The fourth chord generated by Physarum
po lycephalum contained F #4, G4, and A4, which sounded
close to the chord Gsus2/D. However, F #4 and G4 are only
a semi-tone apart and therefore, made the chord sound un-
stable. Hence, F #4 was modified to D5 and thus obtaining
the chord Gsus2/D.
The original vocal tune composed by the author was a
simple melody line. 0% produced a melody that was similar
to the input, but had irregular rhythms and few variations
in pitches. It did not fulfil our goals of making the vocal tune
sound more catchy. 30% produced interesting variations in
the output. However, the output sounded very continuous
to suit a vocal line. Its rhythmic structure did not have
enough stops. 100% offset formed a good foundation for the
vocal line. Furthermore, it went well with the new chords
517
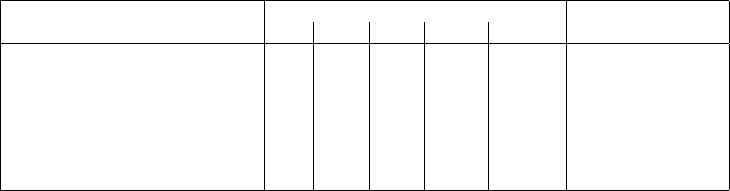
Table 1: Table of tests conduced with PhyBox for different values of measurement offset (MO). Numbers
within the MO columns sh ow the total dissimilarity and ‘-’ indicates no test was conducted. The ‘final song’
column shows the values of MO chosen for the final s ong. The values in bold indicate that they were modified
to suit the song. There are repetitions of values across rows because the dissimilarities are calculated for
the whole block and not only the section.
Section Physarum MO (%) Final Song
0% 30% 50% 100% 100%R
Main Riff 69 214 - 261 - 100%
Verse Pt. 1 Piano 69 214 - 261 - 0%
Verse Pt. 2 Piano - - - - 150 100%R
Pre-chorus Piano - - - - 150 100%R
Piano Solo 414 738 743 860 313 50% and 100%R
Verse Pt. 1 and Pt. 2 Vocals 106 229 - 265 81 100%
Pre-chorus and Chorus Vocals 106 229 - 265 81 -
comp osed by Physarum polyceph alum. However, few notes
were omitted and altered in order t o suit the song.
4.4 Verse Part 2
The rhythmic structure was unaltered because resistors were
connected instead of memristors for time between note-ons
and duration. During the second bar, the memristors pro-
duced an accidental Bb which was not anticipated by the
comp oser. This accidental (or blue note) is pretty popular
in jazz and blues genres, which gave the song a new flavour.
After performing few modifications, the arpeggio composed
by Physarum polycephalum was fit into the song.
4.5 Pre-chorus
The pre-chorus of the song mainly consisted of strumming
and piano patterns in close association with the drumbeat
and hence, we wanted to maintain the rhythmic structure.
For this part of the song, we used resistors for rhythmic pa-
rameters and memristors for pitch and velocity. The m em-
ristors changed the voicing of chords and also performed
some chord inversions. Few modifications were made to in-
corporat e it in the song.
For the vocal tune of the pre-chorus, we felt that the
output of memristors did not match the backing chords.
Furthermore, the original melody line went well with the
subtle changes produced by memristors in t he piano line.
Therefore, we used the original tune.
4.6 Chorus
The chorus of the song had the main riff running in the
background alongside vocals. Therefore, the instrumental
section was a replica of what was done earlier.
In order to compose the vocal tune for the chorus, we
analysed the output of resistors connected to rhythmic pa-
rameters. For the chorus of the song, we felt that the origi-
nal version was more suitable and Physarum polycephalum’s
output did not improve it. As we progressed towards the
chorus, the song was gaining structure and form. Further-
more, it had the main riff of the song playing in the back-
ground. Therefore, there was more rigidity in the number
of suitable musical options. Hence, we chose the original
tune created by the composer.
4.7 Piano Solo
Among all values of measurement offset, 50% was chosen
due to the following explanation. In the original solo com-
p osed by the author, there was a mode shift from E minor
to E Phrygian and then C major. Among all outputs pro-
duced by Physarum polycephalum, 50% created a solo that
was distinctly perceived as G major. Surprisingly, there was
no training data that existed in G major. However, it is the
relative major of the E minor scale. This was a very in-
teresting observation and Physarum polycephalum sounded
like a highly skilled pianist! This demonstrated that the in-
telligence of the organism can be harnessed to create unique
trajectories in creative processes. T here were no modifica-
tions made by the composer on Physarum polycephalum’s
output. Furthermore, 30% offset created few bars that were
p erceived as G major. The output of 30% and 50% were
similar, but 50% was preferred by the composer. 100% offset
created many stops during the solo which made it unsuit-
able. Additionally, 100% offset sounded distinctly different
from 30% and 50%.
The final piano solo starts with the original one comp osed
by the author. This was mutated with outputs produced
by 0% and 100%R. Just before the scale shift to C major,
Physarum polycephalum’s solo in 50% offset begins, which
lasts for 9 bars. In the final song, this part spans from 2
min 22 s to 2 min 42 s.
4.8 Drums
For all values of measurement offset, the musical files sounded
random and unsuitable for the song. This might be due to
the 4 parameters — pitch, velocity, time b etween note-ons,
and duration are not suitable to analyse percussion tracks.
It might need higher-level musical structures such as time
signatures. Furthermore, in such a song, drums are gen-
erally a supporting instrument. Physarum polycephalum’s
output sounded more like a melody line and not a drum-
b eat. Due to the above observations, we decided to not use
Physarum polycephalum for the drum solo.
4.9 Discussion
This pap er presented the process of composing with Physarum
po lycephalum-based m emristors. As mentioned earlier, the
mo del adopted for composition uses note transitions. There-
fore, it do es not account for higher-level musical structures
like time signatures and harmonic functions. Therefore, we
had to make modifications in Physarum polycephalum’s out-
put to incorporate it in the song. It is important to note
that modifications to the output produced by Physarum
po lycephalum was only done in parts where there was a high
involvement of multiple tracks and presence of chords. For
instance, slightly altering piano and vocal lines during the
verse, but not making any modifications in the main riff.
Furthermore, individual tracks were passed separately into
the PhyBox, which may have contributed to this behaviour.
As no changes were made in outputs produced for the
main riff and piano solo, it suggests that the model can be
improved by further understanding higher-level structures.
Examining how instruments interact with each other opens
up possibilities of creating networks of memristors that col-
518
lectively produce a coherent and ready-to-use output.
The author felt that the memristors have changed the
original composition significantly and added its own foot-
print. It was an interactive process where Physarum poly-
cephalum makes suggestions and the composer evaluates
them. However, it is important to not e that many observa-
tions and results were drawn through subjective measures
and aesthetic preferences of the composer.
This paper used different values of measurement offset for
different sections of the song. In figure 3, it can be observed
that 0% captures the value at the top of the spike. The next
sample recorded by the PhyBox is after a steep descent.
Therefore, in order to have more flexibility in measurement
offset, the sampling rate of the system needs to be increased.
We used dissimilarity as the metric to compare differ-
ent values of measurement offsets. This can be improved
by borrowing concepts like entropy from information the-
ory [20, 18]. This will enable us t o explore the statistical
structure of music in greater detail.
5. CONCLUSION
This paper presented a composition that was in collabo-
ration with Physarum polycephalum. It showed how the
non-linear behaviour of memristors can be harnessed for the
pro cess of popular music pro duction. It aims to make UC
technologies reach a wider audience so that non-experts in
UC and musicians can adopt them in the process of music
pro duction. Harnessing Physarum polycephalum has great
p otential in computational creativity and popular music.
We showed that resistors can be creatively incorporated
alongside memristors during musical composition. We demon-
strated that a dwell time of 1 s is suitable for obtaining
the memristor’s non-linear behaviour. This is faster than
an earlier model that used 2 s [6]. Dissimilarity values
show that the measurement offset considerably influenced
the melody produced by the memristor. For each param-
eter, the output created by Physarum polycephalum was
unique. This can be verified with sound files provided in
the database.
In this paper, we trained the model with only one song.
Future work would explore how the system can be scaled
up. This might include creating more complicated learn-
ing models such as networks of memristors. Additionally,
higher-level data structures such as time signatures and har-
monic functions need to be explored.
6. ACKNOWLEDGEMENTS
The authors thank Dr. David Moffat for proofreading t he
article and offering constructive feedback.
7. REFERENCES
[1] A. Adamatzky. Physarum Machines: Computers from
Slime Mould, volume 74 of A. World Scientific, 2010.
[2] A. Adamatzky, J. Jones, R. Mayne, S. Tsuda, and
J. Whiting. Logical gates and circuits implemented in
slime mould. In Advances in Physarum Machines,
pages 37–74. Springer, 2016.
[3] O. Bichler, D. Querlioz, S. J. Thorpe, J.-P. Bourgoin,
and C. Gamrat. Extraction of temporally correlated
features from dynamic vision sensors with
spike-timing-dependent plasticity. Neural Networks,
32:339–348, 2012.
[4] M. Bosi and R. E. Goldberg. Introduction to Digital
Audio Coding and Standards. Kluwer Academic
Publishers, 2003.
[5] E. Braund and E. R. Miranda. On building practical
bio computers for real-world applications: Receptacles
for culturing slime mould memristors and component
standardisation. Journal of Bionic Engineering,
14(1):151–162, 2017.
[6] E. Braund, S. Venkatesh, and E. R. Miranda. Phybox:
A programmable interface for physarum
p olycephalum-based memristors. International
Journal of Unconventional Computing,
14(3-4):217–233, 2019.
[7] L. Chua. Memristor-the missing circuit element. IEEE
Transactions on circuit theory, 18(5):507–519, 1971.
[8] S. Colton, G. A. Wiggins, et al. Computational
creativity: The final frontier? In Ecai, volume 12,
pages 21–26. Montpelier, 2012.
[9] D. Cope. Experiments in Musical I ntelligence. A-R
Editions, Inc, 1996.
[10] J. D. Fern´andez and F. Vico. Ai methods in
algorithmic composition: A comprehensive survey.
Journal of Artificial Intelligence Research,
48:513–582, 2013.
[11] E. Gale, A. Adamatzky, and B. de Lacy Costello.
Slime mould memristors. BioNanoScience, 5(1):1–8,
2015.
[12] K. Jones. Comp o sitional applications of stochastic
pro cesses. Computer Music Journal, 5(2):45–61, 1981.
[13] B. Linares-Barranco and T. Serrano-Gotarredona.
Memristance can explain
spike-time-dependent-plasticity in neural synapses.
Nature Precedings, 4, 09 2010.
[14] E. R. Miranda and J. Al Biles. Evolutionary computer
music. S pringer, 2007.
[15] E. R. Miranda, E. Braund, and S. Venkatesh.
Compos ing with biomemristors: Is biocomputing the
new technology of computer music? Computer Music
Journal, 42(3):28–46, 2018.
[16] T. Nakagaki, H. Yamada, and
´
A. T´oth. Intelligence:
Maze-solving by an amoeboid organism. Nature,
407(6803):470, 2000.
[17] T. Nakagaki, H. Yamada, and A. Toth. Path finding
by tube morphogenesis in an amoeboid organism.
Biophysica l chemistry, 92(1-2):47–52, 2001.
[18] M. T. Pearce and G . A. Wiggins. Auditory
expect ation: the information dynamics of music
p erception and cognition. Topics in cognitive science,
4(4):625–652, 2012.
[19] S. M. Ross. Stochastic processes. JohnWiley & Sons,
Inc, 2 edition, 1996.
[20] C. E. Shannon. A mathematical theory of
communication. Bell system technical journal,
27(3):379–423, 1948.
[21] G. S. Snider. Spike-timing-dependent learning in
memristive nanodevices. In Proceedings of the 2008
IEEE International Symposium on Nanosca le
Architectures, pages 85–92. IEEE Computer Society,
2008.
[22] D. B. Strukov, G. S. Snider, D. R. Stewart, and R. S.
Williams. The missing memristor found. nature,
453(7191):80, 2008.
[23] P. M. Todd. Computer Music Journal, 13(4):27–43,
1989.
[24] S. Venkatesh, E. Braund, and E. R. Miranda. A
nonlinear approach to generate creative data using
physarum polycephalum-based memristors. In
A. Adamatzky, editor, S lime Mould in Arts and
Architecture. River Publishers, 2019.
519
